整理一些资料,对鱼龙的牙齿颌骨和食物做一个概述
海爬食性的判断无非基于骨骼形态,特别是头骨和牙齿形态;以及消化道内容物来判断
关于牙齿,最有效地一般是基于Massare在1987年的研究,一般通过两个方面判断:
(1)齿冠的相对大小(齿冠高度/头骨宽度)(3)冠形指数(齿冠高度/牙齿直径)
三叠纪鱼龙
三叠纪是鱼龙多样性最强的时期,但是鱼形鱼龙却相当少,此时的大部分鱼龙是伏击掠食者,并可能比侏罗纪鱼龙有更广泛的食谱(
1.短尾鱼龙类、混鱼龙类与短头鱼龙类
混鱼龙科包括Barracudasauroides(=茅台混鱼龙Mixosaurus maotaiensis=盘县混鱼龙Mixosaurus panxianensis)、混鱼龙Mixosaurus、Contectopalatus、Phalarodon
短头鱼龙科Omphalosauridae包括短头鱼龙Omphalosaurus、Tholodus、新民鱼龙Xinminosaurus
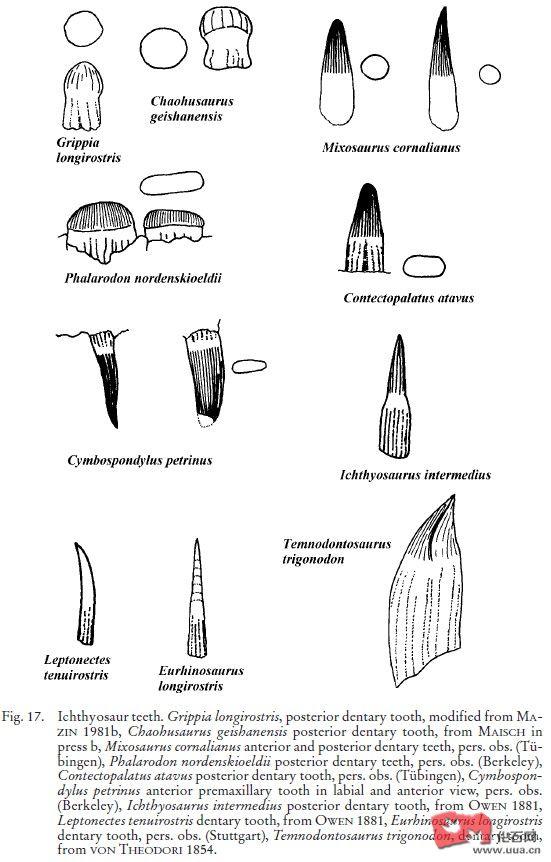
巢湖龙Chaohusaurus的吻较为细长,颌骨后部有一些钝而圆的的牙齿
Barracudasauroides,它显示混鱼龙可能起源于东方
混鱼龙属于混鱼龙亚科Mixosaurinae拥有矢状嵴,一般不超过头骨背部的高度
Mixosaurus cornalianus的后牙较尖,只有几毫米大。而Mixosaurus kuhnschnyderi后牙强壮且钝。
而Phalarodon则属于Phalarodontinae亚科,有非常高的矢状嵴和臼状的后牙,似乎有一个额外的第六指
Phalarodon nordenskioeldii有大的牙齿(2cm),被描述为丘型、球形等形状。颌骨后部有很大的牙齿,比前部的小牙还多;而相对较大的Phalarodon major就没有那么夸张了
Phalarodon fraasi(=Mixosaurus fraasi)的齿冠也有1cm大,类似于亚成体的Mixosaurus nordenskioeldii。它的牙齿有一定的磨损,说明其花了一些时间在破碎坚硬的食物上
其牙齿较为侧扁,也有可能有剪切功能,但是不够锋利。我国也发现过其化石
鱼龙胃内容物一般较少发现,而且有些生物可能有去除猎物硬壳的习惯,所以内容物没有硬甲不能说明猎物就没有硬甲
短尾鱼龙和巢湖龙的牙齿很少有磨损,而且牙齿比较小(巢湖龙牙齿相对大小为0.06),仅有牙齿形状一条显示可能以硬壳生物为食(巢湖龙冠型指数约为1.0),此外短尾鱼龙的牙釉质也较薄,不太可能用来crush,可能只是有时以甲壳类动物为食
而Phalarodon fraasi(可能也包括Mixosaurus nordenskioeldii)则可满足上述要求,它们可能更主要的以硬壳生物为食
不过更专业的贝类捕食者是短头鱼龙科Omphalosauridae,对于其归属,目前还有争议。它们拥有粗短的吻部和好几排不规则排列的牙齿,不像混鱼龙类,它们的牙齿全部由钝牙构成,甚至胜过球齿龙。但是它们和混鱼龙科并没有亲缘关系,球形的牙齿是非同源的。Tholodus和新民鱼龙有完整的化石,它们相当相似
Contectopalatus atavus(=Phalarodon atavus)可能是体型最大的混鱼龙科,为大到特大型,发现于德国Muschelkalk。头长可达70~80厘米,体长至少4,5~5米(SMNS 56315),可能比同一地区的布氏杯椎鱼龙Cymbospondylus buchseri(头长68cm,5~5.5米)还大。事实上,这样巨大的混鱼龙可能不是偶然的现象,Fraas (1891)和Huene (1916)也曾描述过巨大的混鱼龙。它与Phalarodon一样,属于Phalarodontinae,头骨窄,有非常高的矢状嵴供肌肉附着,使得其具有很强大的咬合力。上颌的牙齿较大,下颌的较小,颌骨前部有圆锥形的尖牙来穿刺。后牙侧面有两道很深的沟槽,使得横截面是∞字型,而并不是真正用来破碎用的牙齿(像Phalarodon)。根据Michael和Matzke(1998、2000)的说法,它可能是当地的顶级捕食者
2.贝沙诺龙、萨斯特鱼龙(兼贵州鱼龙)与杯椎鱼龙
贝沙诺龙Besanosaurus目前有一个化石,化石保存长度为5.8米,估计全长5.412米,头长51cm,占体长比例为9.42%,尾长占体长比例为51.47%。其化石旁保存了一只70~80cm长的小混鱼龙
贝沙诺龙拥有非常小的牙齿,最大只有8mm,相对大小也十分小,可能只有0.04。头骨的比例不足10%,拥有细长的吻部,并有非常长的尾巴,显然以小型猎物并很有可能是头足类为食
萨斯特鱼龙属的化石往往较为零碎,这是一属大型鱼龙,可能最大能有7米长,目前包括两个种Shastasaurus pacificus(=Shastasaurus alexandrae、S. altispinus 和S. osmonti)和关岭邓氏萨斯特鱼龙Shastasaurus tangae(=亚洲杯椎鱼龙Cymbospondylus asiaticus 、美丽盘江龙Panjiangsaurus epicharis、(?)卧龙岗卡洛维龙Callawayia wolonggangense)
Shastasaurus pacificus(=Shastasaurus alexandrae)拥有粗壮的吻部,牙齿相对较小。Shastasaurus pacificus(=Shastasaurus altispinus)头长大约1米,接近杯椎鱼龙的长度,体长估计5~6米(不过根据邓氏萨斯特鱼龙的新数据,可能要更长一些)
牙齿最大大约3cm高,齿冠还要少一些,直径1cm多一点(自己测量的),冠形指数大约2左右。根据牙齿形态,Massare(1989)认为它的牙齿属于“cut”的类型,而且根据其又粗又宽的吻部,可能以大型鱼类和其他海爬为食,但更可能是机会主义者,见到什么吃什么
邓氏萨斯特鱼龙荐前椎数极多(65~69),头长最大也有1米多长。牙齿尖锐,向内倾斜,无钝性齿,齿槽外缘高于内缘,应该属于亚槽齿型,所以盘江鱼龙不是靠牙齿磨蚀食物,而是直接吞食。根据研究(潘薪如、江大勇、孙作玉、蔡涛、张大鹏、谢家林2000),其圆锥形的牙齿不够锋利等特征,故不属于凶猛的大型食肉鱼龙。
另外关岭生物群中有10米以上甚至15米的超大型鱼龙,但是还没有仔细研究过。根据陈孝红、程龙2003年的论文,目前定种邓氏萨斯特鱼龙可能仅仅是未成年,而那些10米以上的大鱼龙可能才是成年个体
由于没有一具完全完整的化石,杯椎鱼龙的体长只能估计,Merriam在1908年估计两具帕特杯椎鱼龙的化石组合起来(前半身UCMP 9950 + 后半身UCMP 9947)超过9.1米,而UCMP 9950的头骨长度为1166mm,故估计杯椎鱼龙的头长为体长的13%,布氏杯椎鱼龙的PIMUZ 4351(可能是亚成体)头骨长680mm,估计体长5米(也有可能有5.5米),尼科尔杯椎鱼龙的FMNH PR2251头骨长975mm,估计体长会有7.6米。BBC中的杯椎鱼龙体长达到10米,大部分杯椎鱼龙可能无法达到这一长度。1908年Merriam称,杯椎鱼龙平均体长6.2米(这里当然是指帕特杯椎鱼龙),只有少数大个体才可达9米。有一个头骨显示杯椎鱼龙可达13.5米,如果属实,应该是已命名的鱼龙中第4长的(次于西卡尼秀尼鱼龙Shonisaurus sikanniensis、喜马拉雅鱼龙Himalayasaurus tibetewis和通俗秀尼鱼龙Shonisaurus popularis)其牙齿为异形齿,前牙尖锐而后牙侧扁
3.秀尼鱼龙、喜马拉雅鱼龙
根据Camp于1980年的描述,通俗秀尼较大的头骨(BISP-1)保守估计为2.5米,体长可能有14.5米。而其拥有2.6~3cm的牙齿孔,FZVE-3不超过31.7mm
根据Nicholls和Manabe04年的说法,成年秀尼鱼龙是缺齿的,只有一些非常小的样本具有很小的牙齿。要喂饱这样的大动物是很困难的,Webb (1984)和Alexander (1982)曾建议过滤食的可能性。但是却没有发现已知的滤食结构,而且细长的吻部也不像能够过滤大量的水。基于Massare(1987)的研究,这样的大型鱼龙可能仍然以头足类为主食。其可能像现在的喙鲸一样,通过舌头的抽吸取食,从而不必使用过滤器
而喜马拉雅鱼龙则有大而发达的牙齿,具有切割边缘,与秀尼鱼龙明显不同。它具有很粗的吻部,但是可能只有中等长度(董枝明1972)。根据Motani、Manabe和董老99年的文,它可能以大型脊椎动物为食
回复人:鸿穹神皇上帝
杯椎鱼龙属的脊椎骨数量还是比较多了,Cymbospondylus buchseri保留下来的51块脊椎骨,但是第51块脊椎骨上面的肋骨的长度和前面最长的肋骨相比并没有明显的减短,就此推测其前部Presacral脊椎骨数量应该超过55块,总的来说眼眶小一些的鱼龙估计咬合力要大一些。
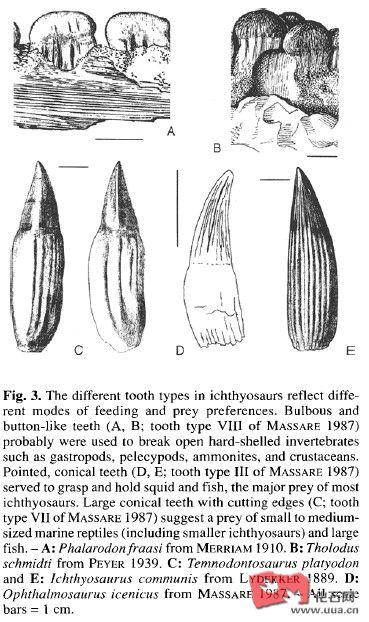
根据Massare87年关于海爬牙齿形态和食性的研究,大部分鱼龙以头足类为主食,拥有4种牙齿Pierce I、Smash、Crunch和Cut,此外,偶尔有一些鱼龙属于Pierce II/General,但三叠纪常见的Crush没有再出现,而且也没有矢状嵴的消息(图39的一些比例可能不适用于鱼龙,图40中黑色的小三角形代表鱼龙,白色圆形为上龙,黑色圆形为蛇颈龙,白色正方形为真蜥鳄,黑色正方形为地蜥鳄)
蛇嘴鱼龙科
Pierce I:蛇嘴龙科往往具有一种细长针状的牙齿,相比较蛇颈龙类中形状类似的牙齿,大小更小且更光滑。冠形指数通常大于3.5,至少不小于3,可能是用来刺穿柔软的猎物用
狭翼鱼龙和大眼鱼龙科
Smash:而包括狭翼鱼龙在内的很多小型后三叠纪鱼龙拥有一种简单的圆锥形牙齿,但有一个圆顶,牙齿的磨损类似于现生抹香鲸,这样的牙齿顶部和磨损显示是用来把持柔软猎物而不是穿刺用的。肠胃内容物也显示它们以头足类为主食,仅有少数鱼类。其冠型指数大约为2.0~3.0,齿冠的相对大小不超过0.15,而且往往小于0.10。事实上有一些鱼龙(包括狭翼鱼龙的模式种Stenopterygius quadriscissus、Chacaicosaurus(属于狭翼鱼龙科)和大眼鱼龙Ophthalmosaurus)牙齿已经发生了退化,减少,甚至无牙。短鳍鱼龙Brachypterygius和扁鳍鱼龙Platypterygius也属于这一类型,但是它们的牙齿发达,更强壮一些,齿冠略有弯曲
幼年泰曼、Suevoleviathan和鱼龙属
Crunch:一些中型的鱼龙(如幼年的板齿泰曼Temnodontosaurus platyodon(=Temnodontosaurus risor)(头长)、Suevoleviathan(4米以上))或一些中小型鱼龙如鱼龙属的模式种Ichthyosaurus communis(一般头长50cm,体长2.5米,最大的头骨发现于瑞士,超过90cm),拥有一种直且粗壮,有粗糙顶尖的牙齿,冠型指数约为2.0~3.0。牙齿表面往往有突出的竖棱,但是不形成切割边缘,旧牙往往有磨光的顶部,显示它们主要以硬的猎物为食,齿冠的相对大小一般小于0.15,显示它们也是以把持为主。其食物来源可能是有内壳的头足类甚至是薄壳的菊石
泰曼鱼龙
Cut:而板齿泰曼Temnodontosaurus platyodon,宽头泰曼Temnodontosaurus eurycephalus和三角齿泰曼Temnodontosaurus trigonodon(=Temnodontosaurus burgundiae、Leptopterygius burgundiae、Leptopterygius acutirostris)这样的大型鱼龙,牙齿则是轻微侧扁、粗壮,有两条锋利的脊作为切割刃,刃的边缘与颌骨长度方向对齐。不像大多数鱼龙,这些牙齿的表面相当光滑,冠型指数约为1.5~2.5。这些牙齿经常有破损,尖端的破损往往是圆而光滑的,三角齿泰曼的腹中曾发现过一只其他的小鱼龙。这些牙齿很类似掠食性沧龙的牙齿,但相对大小却不大。
Aegirosaurus
Pierce II/General:最典型的代表为Aegirosaurus,正模头长56cm,缺了一小块,体长1.77米,可以说头部的比例很大了,最大头长接近1米。牙齿小、多,吻长,齿冠相对大小小于0.1,冠型指数大约为1.7,介于Pierce II(虎鲸也属于Pierce II这一类的)和Smash之间,它的牙齿太小,所以和Smash一样以头足类为食
鱼龙
DONMG:1983.98属于鱼龙属,化石保存于Beechfield House Museum。化石被确认为Ichthyosaurus communis,化石保存了头骨和躯干的前部,包括了腹部地区。化石保存长度为1.1米,其中三分之一是头骨,牙齿的情况符合Massare87年的研究。胃内容物有很多箭石壳的碎片,难以辨认箭石的数目究竟有多少。箭石有四种类型,样本消化道内容物有其中的3种:Type 1,Type 2和Type 3
扁鳍鱼龙
QM F16811是一条7米的雌性Platypterygius longmani(扁鳍鱼龙属),发现于澳大利亚,腹中有一个胎儿(头长40cm)。其胃中发现了protostegid海龟、enantiornithine海鸟和actinopterygian鱼,这些食物没有来得及消化,鱼龙就已死亡。鱼大多是一些2~8mm的椎体,显示个头不大,但有三个大型的牙齿(5~7mm)和一块下颌显示它也会以大型鱼类(大约50cm)为食,可能是鲱状的Cooyoo或Pachyrhizodus。海龟化石为一只或几只幼体,全长10~20cm,估计属于Protostegidae,可能是Notochelone costata。从龟化石的情况看,吞下猎物之前可能有过通过摇晃将猎物撕碎的过程(processed by shake feeding)。海鸟的化石也非常小,跗跖骨只有19.45mm,可能是Nanantius。Kear、Boles和Smith于文中建议双臼椎龙的竞争可能与鱼龙衰落有关(我个人不觉得如此)。
板齿泰曼鱼龙、三角齿泰曼鱼龙、宽头泰曼鱼龙
板齿泰曼和三角齿泰曼是一些瘦长的鱼龙,直线上速度(就比例而言)可能不如后来的狭鳍鱼龙和大眼鱼龙,但是转向的话更方便一些。牙齿齿冠一般只有3cm,但是下颌却往往能有1.5米以上,齿冠的相对大小不足0.10,由此可推知其猎物体型并不大,McGowan在95年的文中认为它们不过是以鱼和头足类为食罢了(不过我个人认为这不是问题,颧突,甚至虎鲸的比例貌似也没有超过0.1么,不过似乎侏罗纪大型海爬都比较青睐头足类?连巨游龙都是……)。泰曼鱼龙的眼眶相对较小,orbital ratio一般小于0.21,且大部分情况下小于0.18。
板齿泰曼的头骨一般比三角齿泰曼来的大,吻部长度中等,但是头部的比例也偏大,因此整体长度差不多,牙齿不是很大,最大58mm(BATGM M3577a,不完整头长1890mm)。板齿泰曼生存于早侏罗纪早期,BMNH R1155是目前最大的头骨,不完整长度超过1.92米,下颌超过2米,并且板齿泰曼还有很多超过1.8米的头骨,最大的完整化石BMNH 2003,头长1.39米,体长至少6.29米。但是头后骨骼的相对大小较小,比如GPIT 1491/13和BMNH 2003的比较,头长相似(1350和1390mm),但是三角齿体长长出一米(5920和4900mm)。英国发现过一个巨大的方骨ANSP 10112,有272mm,还是未定种,但是最有可能是属于板齿泰曼的。而板齿泰曼BMNH R215方骨为200mm(头骨大约1840mm),根据比例估算,有BMNH R215的1.36倍大,通过简单的放大,可以得到一个2.5米的巨大尺寸,,根据BMNH 2003的比例,估计体长11米以上,这个估计是一个很保守的估计,因为泰曼头骨的增长比体型的增长慢。图76为方骨ANSP 10112
相对而言,三角齿泰曼的头骨小得多,连1.6米都勉强。三角齿泰曼生存于早侏罗纪晚期,它的的吻部要长一点,而且鳍也要长一点(指节数28:20)最大的SMNS 50000体长达到9.09米,但头骨只有1.53米(我估计是吻部化石有缺失?),但总的来说1.5米以上的的头骨体长都有个7米多,甚至有一些1.4米不到的记录体长也有7米(我严重怀疑这些头骨的完整性)。Huene 1922年估计三角齿泰曼最大可达11米。
宽头泰曼仅有一个头骨被发现,头长不完整长度950mm,牙齿的相对比例较大,最大额牙齿为54mm,齿冠短,牙齿坚固,颌骨极粗短,95年McGowan的文中被认为和下文所述的未知的大鱼龙一样可能以其他鱼龙为食下图为维基上宽头泰曼的复原,其实复原的有一点错误,泰曼的后肢和前肢大小差的没有那么多
焦点:早侏罗纪巨鱼龙
早侏罗纪有一些零碎的化石记录显示当时的鱼龙可以长到非常大的水平,包括上文提到的巨大方骨、一些大椎体、肩胛骨和大牙齿。由于其巨大的体型和牙齿,McGown建议这些鱼龙以大型猎物为食
泰曼的牙齿最大也不超过6cm,但是记录中最大的牙齿BMNH 28283足有116mm,这个巨大的牙齿有一个很矮的齿冠(33mm),牙根占了牙齿总长的三分之二,有锋利的切割边缘。还有其他一些巨大的牙齿,比如有一个颌骨节片上最大的牙齿ROM 38129,牙高111mm,齿冠38mm。这些牙齿不太可能属于板齿泰曼和三角齿泰曼,因为这二者齿冠的高度往往与齿根长度相差不大(虽然有一些个别的记录)。这些牙齿的外形更接近宽头泰曼,但是大小上却是宽头的2倍多,因此除非宽头正模仅是幼体,否则不可能长到如此之大。
此外还有巨大的肩胛骨和椎体,例如肩胛骨BMNH 481和椎体BMNH R226,,发现于早侏罗纪早期,来自同一样本,同时还发现了这个巨大鱼龙的一些肋骨和椎骨Lydekker (1889)。肩胛骨BMNH 481,有444mm长,而9.1米的三角齿泰曼SMNS 50000肩胛骨不过290mm,仅有前者的65%,按比例放大可以得到一个14米的巨大动物。巨大的椎体BMNH R226,高达205mm,而SMNS 50000仅有124mm,按此比例,这条鱼龙足有15米。而在Banz, Huene1922年的讨论中,有一个个更大的化石样本,包括部分椎骨和肋骨,发现于早侏罗纪晚期,最大的椎体达22cm,甚至略大于BMNH R226,所以可能会有15~16米
补充几种鱼龙的precaudal length/longest rib,计算体重的话用得上
Suevoleviathan disinteger(SMNS 15390) 2.273
Temnodontosaurus trigonodon(SMNS 15950) 2.564
Eurhinosaurus longirostris(GPIT 1025/18/1) 2.5
Stenopterygius quadriscissus(FSF R4154) 1.613
Ophthalmosaurus icenicus(PMAG R340) 1.266
根据狭翼鱼龙2.4米165kg推一下,9米的泰曼大约3.5吨,很瘦了,直接用下面图片的长高比来推也不会超过等长大眼鱼龙的一半。16米大约20吨,不会超过30吨
根据最新的东西的一点补充:
分类调整:
萨斯特鱼龙属Shastasaurus的几个种同物异名
Shastasaurus liangae:
=Guanlingsaurus liangae
Shastasaurus pacificus:
=Shastasaurus alexandrae
=Shastasaurus osmonti
Shastasaurus sikkanniensis:
=Shonisaurus sikkanniensis
萨斯特鱼龙属这三个种的进食现在被认为都是类似于西卡尼的抽吸式
卡洛维龙属Callawayia的几个种的同物异名
Callawayia neoscapularis
=Shastasaurus neoscapularis
Callawayia altispinus
=Shastasaurus altispinus
贵州鱼龙仍然作为一个单独的属,而不是让邓氏贵州鱼龙Guizhouichthyosaurus tangae并入萨斯特鱼龙
贵州鱼龙和卡洛维龙都算是高等的Parvipelvia
可以自己计算得出一些东西
短鳍鱼龙
短鳍鱼龙吻部很粗,牙齿大,被认为咬合力也比较大
其他的测量数据上表中有
对上一楼表格的解释以及一些鱼龙测量方面的东西
Tooth length index可以计算牙齿长度
A. Continuous
1. Orbital ratio: ratio of orbital diameter to jaw length (HIL).
2. Snout ratio: ratio of snout length to jaw length (J/L).
3. Premaxillary ratio: ratio of premaxillary segment to jaw length (I/L).
4. Prenarial ratio: ratio of prenarial segment to to jaw length (B/L).
5. Maxillary tooth count: mean number of maxillary teeth.
6. Postorbital ratio: ratio of postorbital segment to jaw length (D/L).
7. Orbital position index: ratio of distance between centre of orbit and hindmost point of skull to distance between centre of orbit and most anterior point of skull (CIA).
8. Jaw depth ratio: ratio of jaw depth, measured at midpoint of jaw, to jaw length (G/L).
9. Snout depth ratio: ratio of snout depth, measured at midpoint of snout, to jaw length (E/L).
10. Tooth density index: number of upper and lower teeth counted in 10% of jaw length,centred at midpoint of snout (F).
11. Tooth length index: 10 x crown length of longest tooth divided by jaw length.
B. Multistate
12. External naris shape: petaloid (Fig. 5A), markedly elongate (Fig. 5B), or curviform (Fig. 5C).
13. Extracondylar extent: Extracondylar area (6) is much less extensive than condylar area (4) (Fig. 2A); areas are approximately equal (Fig. 2D), or extracondylar area is much more extensive than condylar area.
C. Dichotomous
14. Skull profile: rostrum grades gently into cranium (Fig. 5D) or is demarcated from cranium (Fig. 5E).
15. Condylar differentiation : basioccipital condyle is clearly defined and set off from extracondylar area (Fig. 2D) or is not (Fig. 2A).
16. Position of stapedial facet: stapedial facet (3) of basioccipital extends ventrally (Fig. 2D), or does not (Fig. 2A).
17. Basioccipital profile: anterior surface (5) of basioccipital is flat (fig. 2B), or convex (Fig. 2E).
18. Basioccipital peg: anterior surface of basioccipital is often extended into a peg (McGowan 1973a, p. 12, fig. 2). Peg (7) is well developed (Fig. 2E), weakly developed, or absent (Fig. 2B).
(本文为化石网论坛精华帖,由网友蓝藻脊提供,仅供大家学习参考。编辑:常君滢)