

根据一些资料显示 , 从1580年至2008年间袭击人最多的十大鲨鱼中 , 大白鲨是首居第一 , 攻击人的记录达377次,致死人数有205人;居氏鼬鲨是居第二 , 攻击人的记录达137次,致死人数有82人 ; 公牛鲨攻击人的记录有90次,致死人数有64人。在第二次世界大战间 , Nova Scotia号轮船在南非附近被德军潜艇用鱼雷击沉 , 1000名乘客只余192名生还者 ; 1945年7月30日,有最少60至80名水手被鲨鱼袭击至死 , 二次鲨类袭击人类的事件都被认为和白鳍真鲨有关 。
真鲨科虽没有鼠鲨科般具有巨大的体型,但它们的凶猛程度绝不亚于鼠鲨科 , 尽管真鲨目也没有像鼠鲨目般有过称雄第三纪海洋的光辉岁月 , 但它们经历漫长的岁月还能维持种类的多样性 , 而且也有新属种被发现 , 例如光齿真鲨 , 南非副沙条鲨(Paragaleus leucolomatus)及澳大利亚半沙条鲨(Hemigaleus australiensis)就是真鲨目的新种。 相反鼠鲨目像黄昏的太阳 , 经历了第三纪灿烂光辉的黄金岁月后 , 支持至现在不过只有16种 , 从多样性来看二者形成了强烈的对比。每人对强者的理解各有不同 , 有人会认为有过称雄海洋的鼠鲨科及黑鲸类是强者 , 但也有人会认为幸存才是强者 , 现存真鲨目中有不少成员早于第三纪时期已存在于地球上 ,有些种类甚至比巨齿鲨还要早出现 , 大西洋斜锯牙鲨于始新世时期己存在了。真鲨目成员遍布浅海 , 远洋 , 深海 , 泻湖 ,红树林,珊瑚礁甚至是淡水 , 它们有底栖性 , 大洋性及淡水性等各种形式, 例如猫鲨属种就是偏好多沙海床上的底栖性鲨类 ; 东太平洋就能看到数量上百的路氏双髻鲨在成群游动 , 大青鲨在大西洋随强大的洋流追捕鱼类及乌贼 ;黑边拟皱唇鲨(Pseudotriakis acrages)及小齿拟皱唇鲨(Pseudotriakis microdon)就能在1500公尺的水深生活 ; 柠檬鲨就是在红树林及泻湖繁殖的 ; 公牛鲨的调节盐度能力比任何鲨鱼都出色 , 它们会定期出现在河流及湖泊 , 在密西西比河及亚马逊河中 , 两处的公牛鲨更能逆流超过2500公厘到达目的地 , 此外巴基斯坦 ,印度及澳大利亚的河流也有露齿鲨属在那里长期生活 。只要是有水的地方 ,就没什么是它们不能征服的 , 能幸存下来并发展出多样性的种类 , 真鲨目才是海洋中真正的强者!
真鲨目是起源于晚侏罗纪时期 , 这和鼠鲨目起源的时间相符 , 最早期的鼠鲨目成员是古锥齿鲨属Paleocarcharias。像翼手须鲨属Pteroscyllium牙齿同时具备鼠鲨科及猫鲨科的特征 , 它们是生活于浅海泻湖的鲨类。斯皮顿翼手须鲨Pteroscyllium speetonensis 牙齿是被发现于英国斯皮顿阿普第阶(aptian)地层 , 牙齿高度达3.5mm , 斯皮顿翼手须鲨Pteroscyllium speetonensis牙齿形状像锥齿鲨类,有直立的主齿尖及一对发展完善的侧齿尖 , 有“V”形的牙根 , 斯皮顿翼手须鲨Pteroscyllium speetonensis有一定的异型齿系 , 细小的牙齿有较少量的唇面脊 , 后牙有不对称的牙根裂(基部)及短小的主齿尖 , 后牙有直立的主齿尖及一对等尺寸的侧齿尖并和主齿尖相平行。翼手须鲨属Pteroscyllium除了斯皮顿翼手须鲨Pteroscyllium speetonensis,还有英国斯皮顿阿普第阶(aptian)地层发现的Pteroscyllium ornatum , 英国晚白垩纪科尼亚克阶(Coniacian)至坎帕阶(Campanian)地层发现的Pteroscyllium hermani , 桑托阶(Santonian) 黎巴嫩的Pteroscyllium signeuxi, 德国森诺曼阶地层发现的Pteroscyllium nolfi及麦斯特里希特阶(Maastrichtian)摩洛哥的Pteroscyllium lamranii 。英国晚白垩纪科尼亚克阶(Coniacian)至坎帕阶(Campanian)地层发现的Pteroscyllium hermani和德国森诺曼阶地层发现的Pteroscyllium nolfi关系较密切 , 有细长的牙齿,纤弱的侧齿尖及 不规则的纹状,牙齿唇面长有直脊; Pteroscyllium ornatum于翼手须鲨属种中有较大的牙齿 , 牙齿高度能达4.5 mm , 有粗状及锋利的主齿尖 , 牙齿主齿尖及侧齿尖也长有锯齿切边。 翼手须鲨属Pteroscyllium和其他猫鲨科的关系至今不明 , 翼手须鲨属Pteroscyllium牙齿有血管并有着其他猫鲨科的齿系模式 , 但它们的牙齿形态却像鼠鲨目 , 翼手须鲨属Pteroscyllium的发现显示了鼠鲨目和真鲨目于进化枝上是属于姐妹群。
德国晚森诺曼阶地层发现的鲨类牙齿化石 :
1. Ptychodus decurrens; 2. Chiloscyllium greenei; 3. Lamna appendiculata; 4. Lamna arcuata; 5-6. Isurus denticulatus ; 7. Scyliorhinus aff. destombesi ; 8. Pteroscyllium nolfi; 9. Squalicorax falcatus; 10. Heterodontus canaliculatus; 11. Hexanchus microdon.
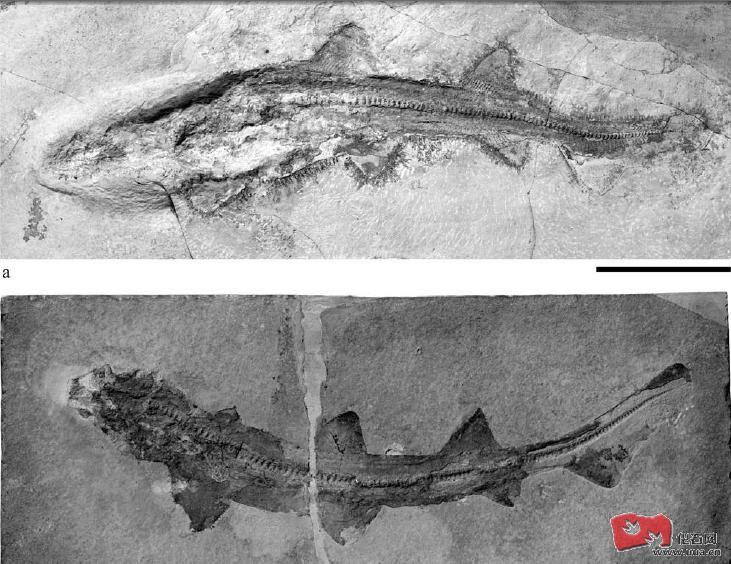
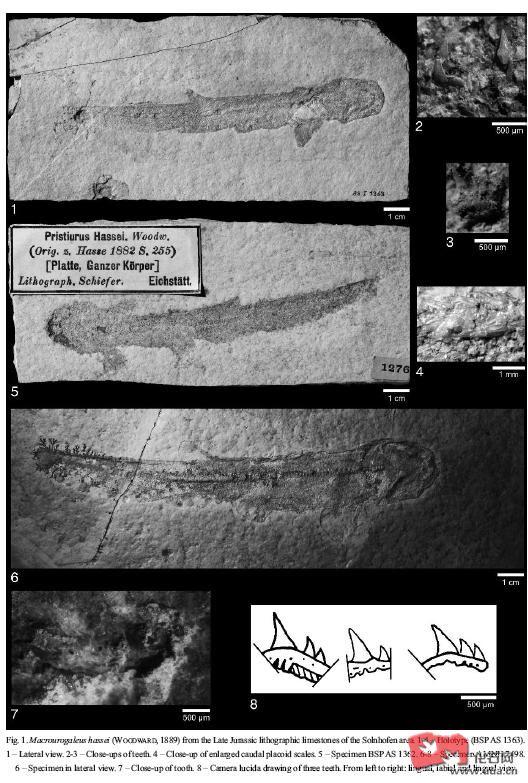
猫鲨科(Scyliorhinidae)是真鲨目中最古老的成员 , 已知最早期的成员来自晚侏罗纪时期包括法国的栉猫鲨Ctenascyllium kimmeridgienus及德国的Macrourogaleus hassei , Macrourogaleus hassei有单一的背鳍,背鳍及臀鳍的位置及大小也不相同, Macrourogaleus hassei有较长的身躯。科律司鲨Corysodon ceirinensis是另一种发现于德国西南部的鲨类,它和古须鲨属Palaeoscyllium有着相似位置及大小的背鳍, 科律司鲨Corysodon ceirinensis化石保存的相当清晰可见, 科律司鲨Corysodon ceirinensis曾被认为和古猫鲨属Palaeoscyllium是同义但至今分类位置还是不明的。相比科律司鲨Corysodon ceirinensis, 古须鲨属Palaeoscyllium formosum 的第二背鳍要比前背鳍的小, 腹鳍及臀鳍位置也各不相同, 古须鲨属Palaeoscyllium formosum牙齿具备精巧脆弱的中齿尖,侧齿尖具有许多雕纹。古须鲨属的Palaeoscyllium formosum分布于德国及西班牙, Palaeoscyllium formosum的相关近亲有英国侏罗纪中期巴通阶(Bathonian)地层发现的Palaeoscyllium tenuidens , Palaeoscyllium tenuidens是最古老的古须鲨属种,于早白垩纪阿尔布阶(Albian) 的英国还有Palaeoscyllium reticularis, 不过Palaeoscyllium reticularis则可能是划作别属Parasymbolus。英国晚白垩纪科尼亚克阶(Coniacian)至坎帕阶(Campanian)地层发现的Palaeoscyllium striatum 前颌的侧齿较大并弯曲及有较细小的侧齿尖 , 齿冠基部长有脊纹。
中生代的英国对真鲨目来说可谓是流奶与蜜之地 , 在当地也发现了许多的真鲨目成员的牙齿化石 , 在这贴会首先论述英国的真鲨目属种。
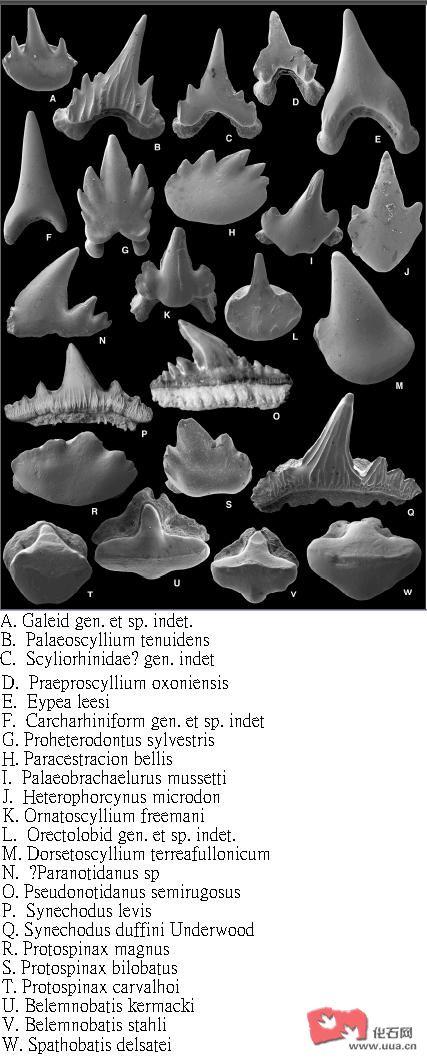
于英国中部及南部侏罗纪巴通阶(Bathonian)沉积层发现了数目众多的鲨鱼牙齿化石 , 牙齿化石多达8000颗 , 当地的鲨类超过25种 , 其中有不少是属于真鲨类 , 发现的软骨鱼类品种如下: 古须鲨属种 Palaeoscyllium tenuidens., Praeproscyllium oxoniensis., Eypea leesi., 原虎鲨属种 Proheterodontus sylvestris., 副异齿鲨属种 Paracestracion bellis., 古长须鲨属种 Palaeobrachaelurus mussetti., Heterophorcynus microdon., Dorsetoscyllium terraefullonicum., Ornatoscyllium freemani., 伪南喙哈那鲨属种 Pseudonotidanus semirugosus, Synechodus duffini, Protospinax magnus , Protospinax bilobatus., Protospinax carvalhoi., Belemnobatis kermacki , Belemnobatis stahli , Spathobatis delsatei , Paranotidanus, 弓鲨属种Hybodus levis及 Breviacanthus brevis等等 , 这些软骨鱼类各自局限于不同的生态环境 , 当时的英国正是被一片浅海所覆盖。
在当地较常见的真鲨目成员是Eypea leesi , 它的牙齿非常普遍 , 有较大的牙齿。Eypea leesi牙齿化石被发现于混浊的离岸相环境及碳酸石灰岩 , Eypea leesi牙齿化石于泻湖及浅水沉积层是没有发现的 , 这显示Eypea leesi是一种偏好深海的鲨类。Eypea leesi的后齿有较低的齿尖 , 这显示它是一类偏好以甲壳类为食的真鲨类。另一种真鲨目成员古须鲨属种Palaeoscyllium tenuidens则和Eypea leesi不相同 , Palaeoscyllium tenuidens则是较偏好浅水的真鲨类。当地的真鲨目成员还有Praeproscyllium oxoniensi牙齿化石于当地的泻湖沉积层非常常见 ,那里的盐度较低 , Praeproscyllium oxoniens是非常偏好浅水的鲨类 。于当地发现的Dorsetoscyllium terraefullonicum, Ornatoscyllium freemani, Heterophorcynus microdon及古长须鲨属种Palaeobrachaelurus mussetti是属于须鲨目成员 ,四种须鲨目成员都是较偏好浅水的鲨类。
于英国晚白垩纪科尼亚克阶(Coniacian)至坎帕阶(Campanian)地层也出土了大量鲨类及鳐类的牙齿化石 , 其中有不少是属于真鲨目成员包括伪猫鲨属种Pseudoscyliorhinus reussi , 厚齿猫鲨Crassescyliorhinus germanicus , 猫鲨属种Scyliorhinus elongatus, Scyliorhinus brumarivulensis., 古猫鲨属种Palaeoscyllium sp., 原宽瓣鲨Prohaploblepharus riegrafi, 栉猫鲨属种Cretascyliorhinus sp., 二个猫鲨科不定种(Scyliorhinidae incertae sedis), 翼手须鲨属种Pteroscyllium hermani , 原猫鲨属未定种Protoscyliorhinus sp., 白垩须狗鲛Leptocharias cretaceous , Palaeogaleus havreensis , 副皱唇鲨属种Paratriakis subserratus , Paratriakis tenuis sp , Paratriakis sp. indet. 及弯齿鲨属未定种(Loxodon sp) 。这些白垩纪鲨鱼属种分别归属原鲨科(Proscylliidae) , 须狗鲛科(Leptochariidae)及真鲨科(Carcharhinidae) , 显示真鲨目成员于当地是非常繁盛。
伪猫鲨属Pseudoscyliorhinus属种包括森诺曼阶(Cenomanian)德国的Pseudoscyliorhinus schwarzhansi及桑托阶(Santonian)至坎帕阶(Campanian)英国的Pseudoscyliorhinus reussi。Pseudoscyliorhinus reussi 有较平坦的齿根及少量的营养构 , 前齿唇面基部处有网状的的纹。伪猫鲨属Pseudoscyliorhinus的牙齿和其他猫鲨科属种很不相同,但是Pseudoscyliorhinus reussi曾被划作猫鲨属种Scyliorhinus reussi 。现存的猫鲨属种幼体有较大的牙根 , 幼猫鲨属牙根也较成体的来得平坦 , 因此发现的Pseudoscyliorhinus reussi牙齿化石也可能是来自幼鲨的 , 发现的伪猫鲨属Pseudoscyliorhinus reussi牙齿化石宽度仅只有3毫米。
Crassescyliorhinus学名有厚的意思 , 故名思义 , Crassescyliorhinus牙齿较其他猫鲨科成员的牙齿来得厚,因此学名也可翻译作厚齿猫鲨。厚齿猫鲨属Crassescyliorhinus germanicus有非常对称的粗实牙齿 , 中央的主齿尖呈圆锥形并占了主齿冠很大的面积比例 , 牙根没有营养构 , 齿冠表面存有脊纹。厚齿猫鲨Crassescyliorhinus germanicus和桑托阶(Santonian)至马斯特里赫特期(Maastrichtian)猫鲨属种Scyliorhinus musteliformis关系密切 , Scyliorhinus musteliformis可能也是归属厚齿猫鲨属,如果真的划归的话,那厚齿猫鲨属Crassescyliorhinus则一直延存至晚白垩纪的马斯特里赫特期(Maastrichtian)。
英国东部白垩纪阿尔必阶(Albian)晚期的Hunstanton Formation发现了另一类猫鲨科属种Parasymbolus reticularis的牙齿化石 , 被发现一共有五块牙齿化石 , Parasymbolus reticularis齿冠唇面上长有网状的纹。Parasymbolus属的主模式种是晚侏罗纪法国的Parasymbolus octevillensis , Parasymbolus octevillensis及Parasymbolus reticularis齿冠及牙根形态相似 ,齿冠唇面长有锋利的脊 , 但不同在于Parasymbolus reticularis齿冠唇面长有网状的纹 , Parasymbolus reticularis牙根的营养构和Parasymbolus octevillensis幼鲨的相似 。Parasymbolus reticularis的发现显示了Parasymbolus是很长寿的属 , 属种生存年代是侏罗纪早期至白垩纪早期。
英国晚白垩纪科尼亚克阶(Coniacian)至坎帕阶(Campanian)发现的原宽瓣鲨属种(Prohaploblepharus sp)的牙齿就非常类似现在的埃氏宽瓣鲨(Haploblepharus edwardsii) , 不过原宽瓣鲨属种(Prohaploblepharus sp)有较直的主齿尖,齿尖高度值要较宽度值来得大 , 且长有两对分散的侧齿尖 , 齿冠基处也长有纵向的脊纹。原宽瓣鲨属(Prohaploblepharus)的主模式种是坎帕阶(Campanian)德国的Prohaploblepharus riegrafi , 牙齿的纹和英国晚白垩纪科尼亚克阶(Coniacian)至坎帕阶(Campanian)地层发现的史前猫鲨属种Scyliorhinus brumarivulensis非常相似 , 不过原宽瓣鲨属种Prohaploblepharus riegrafi有较大的牙齿 , 牙根也有所不同。
和鼠鲨目相反 , 猫鲨科于始新世的普里阿邦阶(Priabonian )的乌拉尔山脉只是属于地域性 , 当地主要的猫鲨科只有三种 , 包括Premontreia uralica , Foumtizia zhelezkoi 及Foumtizia pattersoni 。Foumtizia属的主模式种是Foumtizia abdouni , 这属是小型猫鲨科属系,它们的牙齿相当细小但具有尖锐的齿冠 , 有发展完善的突出齿尖 , 齿冠唇面下部膨大并被珐琅质覆盖及悬垂于齿根之上 , 齿根有发育良好的营养构。Foumtizia abdouni生活于古新世塔内提阶(Thanetian)的摩洛哥 , 其它属种包括古新世达宁阶(Danian)摩洛哥的Foumtizia gadaensis 及Foumtizia arba ,始新世伊普雷斯阶(Ypresian)英国的Foumtizia pattersoni。于西伯利亚西部出土了无数鲨类的牙齿 , 其中也有真鲨目中的猫鲨科牙齿化石发现 , Premontreia uralica., Foumtizia zhelezkoi 及Foumtizia pattersoni 就是被发现于西伯利亚西部盆地的三种猫鲨科成员 , 三种猫鲨科成员牙齿化石的发现显示了它们是于晚始新世时期海洋退减时停留在Peri-Tethys,该地北部于普里阿邦阶(Priabonian)气候冷却作用促进了当地物种形成。
Premontreia是一类已灭绝的猫鲨科属种 , 大多是生存于早古新世至早始新世 , 它们有相对较大的齿尖 , 高度能达6 mm , 齿舌面平滑及外侧突起 , 齿唇面则没有像舌面般有较明显的突面 , 基部有珐琅脊(背褶叠), 牙齿长有一至二对侧齿尖 , 牙齿有膨大的齿根及有显著直立的齿冠。Premontreia属包括达宁阶(Danian)至始新世伊普雷斯阶(Ypresian)前期摩洛哥的Premontreia subulidens , 晚古新世塔内提阶(Thanetian)摩洛哥的Premontreia peypouqueti , 始新世伊普雷斯阶(Ypresian)晚期法国的Premontreia degremonti , 始新世普里阿邦阶(Priabonian)俄国的Premontreia uralica。始新世伊普雷斯阶晚期法国的Premontreia degremonti化石是较常见的 , 于维吉尼亚始新世伊普雷斯阶晚期的Nanjemoy Formation也发现了Premontreia degremonti相似种Premontreia cf degremonti的牙齿化石 , Premontreia属种还有古新世塔内提阶(Thanetian)的法国北部 , 始新世的比利时 , 法国及美国的Premontreia gilberti及渐新世至上新世欧洲的Premontreia dachiardi。近来于比利时始新世路德阶地层也发现了新种Premontreia lutetiensis , 它的牙齿高度达8毫米 , 它的牙齿是Premontreia属中最大的 , Premontreia degremonti牙齿高度也不过才只有6毫米。
光尾鲨属(Apristurus)一共至少有25种 , 大多数体型都是非常细小 , 像圆鳍光尾鲨(Apristurus spongiceps)全长才只有50厘米 , 巴拿马光尾鲨(Apristurus stenseni)全长只是超过23厘米 , 但是属中的驼背光尾鲨(Apristurus gibbosus)雄鲨可达3.88米, 雌鲨更能达4.1米。一些种类如无斑光尾鲨 Apristurus acanutus甚至能栖息于840米的水深 , 南非光尾鲨(Apristurus saldanha)的栖息深度也能达915米 , 北美的加州光尾鲨(Apristurus kampae)及深水光尾鲨(Apristurus profundorum)更能栖息于1800米水深的环境。 不像猫鲨属Scyliorhinus, 光尾鲨属(Apristurus)的化石记录相对要稀少得多 , 已知的史前的光尾鲨属仅只有法国始新世地层发现的Apristurus sereti , 于意大利Fiumefreddo更新世时期沉积层发现了冰岛光尾鲨的亲近种Apristurus aff laurussonii的牙齿化石 , 于当地也发现了黑口锯尾鲨( Galeus melastomus)的牙齿化石。
锯尾鲨属(Galeus)一共约十二种, 于法国的早期中新世地层留下最古老的化石记录,像现存的黑口锯尾鲨( Galeus melastomus)于中新世的法国及德国均有化石记录,已灭绝的成员还有Galeus pristodontus , 不过据说墨西哥始新世中期地层也有锯尾鲨属未定种的牙齿化石发现。梅花鲨属(Halaelurus)及绒毛鲨属(Cephaloscyllium)于印太平洋有广泛的分布 , 其中澳洲也有两属的分布 , 梅花鲨属(Halaelurus)及绒毛鲨属(Cephaloscyllium)都各有一种生活于东太平洋 , 两个属于大西洋都没有分布。绒毛鲨是喜欢栖居于海床石缝及大型海藻,受到惊吓时会吞下水或空气使身体膨胀到正常身躯的两倍大左右 ,使敌人难以把它拉离移居点 。于加利褔亚Kern County也发现了绒毛鲨属未定种的化石。
新软骨鱼类Neoselachians首次于古生代出现 , 包括所有现存的鲨类及鳐类 , 牠们的牙齿由三层珐琅质覆盖。除了鲨类和鳐类外 , 还包括马克默多鲨Mcmurdodus , 吉特鲨Ginteria, 哥利鲨Cooleyella ,Reifia , Hueneichthys, Rhomphaiodon ,小弓鲨"Hybodus" minor, 哥臣鲨Grozonodon candaui , 晚三迭纪至早侏罗纪西欧的Jurabatos cappettai , 早侏罗纪意大利的Ostenoselache stenosoma , 晚白垩纪德国的副扁鲨Parasquatina cappetai及始新世的Odontorhytis 等许多史前软骨鱼类 ; 真正的鲨类除了现存的九个鲨目外 , 还有已灭绝的古鳍棘鲨目(Synechodontiformes) , 古鳍棘鲨目包括原鳍棘鲨属Protospinax, 楔齿鲨属Sphenodus, 古鳍棘鲨属Synechodus, 副楔齿鲨属Paraorthacodus , Welcommia , 至于伪南喙哈那鲨属Pseudonotidanus是否归属古鳍棘鲨目或者是归属六鳃鲨目则尚存疑问 , 于以下并不介绍伪南喙哈那鲨属Pseudonotidanus。
最早期已知的新软骨鱼类成员是马克默多鲨Mcmurdodus , 生存于泥盆纪中期(距今三亿九千万年)的西昆士兰。今人震惊的是 , 这种鲨类较裂口鲨Cladoselache及南极鼠鲨Antarctilamna更符合现在所有鲨目成员的先祖 , 马克默多鲨Mcmurdodus牙齿珐琅质有现在鲨鱼牙齿般特质 , 现存的鲨类有多层的齿排及不同的牙齿形式 , 这特征于大多数远古鲨类是没有 ,但异棘鲨类Xenacanthus却是例如的。如果马克默多鲨Mcmurdodu , 裂口鲨Cladoselache及南极鼠鲨Antarctilamna是同一时期的世系成员的话 , 那么泥盆纪早期已进行了很巨大的软骨鱼类大幅射进化。尽管裂口鲨Cladoselache , 南极鼠鲨Antarctilamna及其它早期软骨鱼类坚持了一段时间,但只有马克默多鲨Mcmurdodus完成了一个伟大的进化梦想。
马克默多鲨Mcmurdodus和现在鲨类关系怎样是相当不明,像大多数早期的鲨类般, 马克默多鲨Mcmurdodus也仅只留下少量的牙齿化石, 牠们的下颌齿像现存的六鳃鲨属和竺鳞鲛属 , 不过最后的马克默多鲨Mcmurdodus和最早的六鳃鲨科成员是存有一亿九千万年的空隙 , 但马克默多鲨Mcmurdodus作为最早的新软骨鱼类成员已是被公认的事实。
吉特鲨Ginteria fungiforma是生存于石炭纪早期的英国及俄罗斯的板鳃类 , 牠们的牙齿非常细小独特 , 牙齿最高值只有0.54 mm , 齿冠像一个台座 。吉特鲨Ginteria fungiforma 齿冠底部长有数目众多的小孔可能是存有单一的血管连接齿冠底部及顶部。另一类软骨鱼类哥利鲨属Cooleyella也有类似的牙齿 , 但吉特鲨Ginteria fungiforma的齿冠表面要较哥利鲨属Cooleyella更呈凹面 , 相比较下吉特鲨Ginteria fungiforma牙齿基部表面较平坦 , 而哥利鲨属Cooleyella牙齿要较吉特鲨Ginteria fungiforma的大得多。
Duffin 及Ward 就于英格兰及威尔士石炭纪前期地层及美国内华逹州二叠纪早期地层发现的牙齿化石建立了一个新科Anachronistidae , 主模式种是Anachronistes fordi 。但后来Duffin, Richter 及Neis宣称Anachronistes是和哥利鲨属Cooleyella为同物异名。哥利鲨属种Cooleyella fordi有三十五颗牙齿 , 牙齿长度介乎于1亳米至2亳米 , 牙齿化石是被发现于北韦尔斯的克卢伊达郡 , 牙齿有圆锥形的主齿尖 , 并有发育良好的侧齿及基部突缘 , 牙根呈V状 , 牙根是布满血管。哥利鲨属Cooleyella有着扁鲨属Squatina及须鲨属Orectolobus的特征如牙齿根部中央有着布满血管的小孔 , 正视时哥利鲨属Cooleyella牙齿排列明显有数行 , 这点于扁鲨属Squatina是存在的 , 齿片间的牙齿齿侧互相重叠 , 哥利鲨属Cooleyella牙齿是被认为适合以压碎甲壳类为食且哥利鲨属Cooleyella是一类底栖性的软骨鱼类。Duffin及 Ward认为哥利鲨属Cooleyella可能像现在的扁鲨属Squatina及须鲨属Orectolobus有着较扁平的身体及较大的胸鳍 。
新软骨鱼类Neoselachians于三叠纪时期繁洐 , Mucrovenator和相关属种小弓鲨"Hybodus" minor , Rhomphaiodon是当时的代表 , 牠们的牙齿形态有着晩三叠纪时期的古鳍棘鲨目Synechodontiformes成员特征 , 并可能也归属于古鳍棘鲨目Synechodontiformes 。 古鳍棘鲨目Synechodontiformes 中的古鳍棘鲨属Synechodus数个种类是生存于三叠纪时期 , 牙齿形态是低且具有较大的齿根 , 齿根布满不规则的小孔 , 有单一或多直立的尖峰 , 这些特征于其它三叠纪的种类如哥臣鲨Grozonodon及伪姥鲨Pseudocetorhinus是具有, 伪姥鲨Pseudocetorhinus是一种三叠纪的滤食者鲨类。
尽管这些三叠纪古鳍棘鲨目Synechodontiformes种类显微结构上有些特征是不相同,但彼此间的关系已是非常密切 。1977年发现的Hueneichthyes , 对这种鲨鱼的认知还是非常少 , 但于1999年已被Cuny及 Benton视作可能是归属古鳍棘鲨目synechodontiform。Hueneichthyes是晚三迭纪的软骨鱼 , 但牙齿形态却有别于其它三迭纪的新软骨鱼类(neoselachians) , Hueneichthyes 有着细长的牙根及少量的小孔 , 平坦的齿冠悬垂于齿根之上。
1980年发现晚三叠纪的Reifia minuta, 牠的牙齿也是不同其它三叠纪的新软骨鱼类neoselachians , Reifia minuta有着平坦的齿冠及纤细的齿根 , 齿根长有数目较少的小孔。形态学上的独特之处显示新软骨鱼类Neoselachii于三叠纪中期或早晚三叠纪时期发生辐射性的分枝演化 , 1982年发现的Vallisia 及1978年发现的伪铠鲨Pseudodalatias 牙齿形态和新软骨鱼类neoselachians一些成员相似但没有典型的新软骨鱼类neoselachians显微结构 , 牠们的关系仍是不明的。
古棘鳍鲨目Synechodontiformes成员有着相同的牙齿形态 , 特点是包括有较低及较大的牙根 , 牙齿有单一及多齿尖 , 牙根上满布不规则排列的小孔 , 三迭纪的哥臣鲨Grozonodon 及伪姥鲨Pseudocetorhinus 并不属于古棘鳍鲨目 , 但却有着以上特征仅只是牙齿中微观结构上的分别。
有关晚三叠纪时期的古鳍棘鲨目Synechodontiformes及六鳃鲨目(Hexanchiformes) , 古鳍棘鲨属Synechodus及其它古鳍棘鲨类的关系是被视作存有疑问的 , Maisey (1985) , Maisey (2004) 及Cappetta (1987)认为牠们不能形成一个单元的类群。古鳍棘鲨目Synechodontiformes看似缺少许多起源的先祖特征而且牠们也可能是并系(paraphyletic) , 牠们的齿式许多特征具有和六鳃鲨目相似性 , 例如牙齿上都具有多尖峰 , 有对称的联合齿 , 特化具压碎特征的后外侧牙齿 , Welcommia及伪南喙哈那鲨Pseudonotidanus二属鲨类形态就介乎于古鳍棘鲨目Synechodontiformes及六鳃鲨目之间。尽管有着这些相似性 , 所有古鳍棘鲨目Synechodontiformes成员有着星状椎骨(asterospondylous vertebra) , 和六鳃鲨目的单环椎(cyclospondylous vertebra)明显不同 。尽管椎骨上的不同显示两个鲨目之间的关系疏远 , 但中生代的六鳃鲨目Hexanchiformes成员具有星状椎骨(asterospondylous vertebra)且椎骨是有纵向的薄层 , 这显示较古鳍棘鲨目Synechodontiformes来得晩出现的六鳃鲨目单环椎结构是具祖先特征并是一种过渡的型态。
于三叠纪晚期的浅海海床生活了一种小型鲨类---小弓鲨'Hybodus minor , 牠的牙齿是平坦且宽阔,显示牠们的牙齿是以压碎食物为主的 ,故名思义牠们是属于弓鲨科, 但最近对牙齿的研究显示牠们可能是归属新软骨鱼类Neoselachii 。于发现小弓鲨'Hybodus minor牙齿化石的地方同时也发现了新软骨鱼类(Neoselachii)另一品种Nemacanthus monolifer的棘部 , 这些鲨鱼没有完整化石被发现 , 因此最初也难显示化石是否属于同一物种 , 直至小弓鲨有完整的化石被发现才可解答鲨类间的关系。Nemacanthus monolifer要比古鳍棘鲨属[Palaeospinax]要来得早 , 牠和古棘鲨属[Palaeospinax]有密切的关系 , Nemacanthus monolifer也被认为有可能是古棘鲨属的先祖, Cappetta于1987年把Nemacanthus monolifer划作古棘鲨科(Palaeospinacidae) , 他们认为Nemacanthus monolifer是真正的新软骨鱼类或者是属于新软骨鱼类的姐妹群。
Rhomphaiodon nicolensis被发现于法国三叠纪晚期地层, 牠有百多颗牙齿化石, 从形态上Rhomphaiodon nicolensis牙齿和小弓鲨"Hybodus" minor 的相似 ; 另一种于法国东部Grozon发现的新软骨鱼类哥臣鲨Grozonodon candaui的牙齿珐琅质结构和小弓鲨的相同 , 二者可能存有密切关系。
Reifia minuta 仅只有五颗细小的牙齿化石发现 , 化石发现于德国诺利克阶(Norian)前期地层 , Duffin (1980)从Reifia minuta 的牙齿判断牠们和须鲨目(Orectolobiformes)关系较密切 , 但是Cappetta (1987)则为认为Reifia minuta 和真鲨目关系要较密切一些 , 但是也有学者认为Reifia minuta的牙齿和锯鳐目中Sclerorhynchidae的牙齿相似。
相对来说大多数新软骨鱼(neoselachian)体型细小 , 长度维持于1米内 , 但牠们的牙齿比弓鲨目来得进步 , 具备了抓捕猎物的能力了 , 例如雷蒂亚古鳍棘鲨Synechodus rhaeticus , 牠的牙齿同时具有抓捕及压碎的能力 ,不过牠们是倾向以甲壳类为食的。当地的另一种新软骨鱼(neoselachian)伪姥鲨Pseudocetorhinus pickfordi可能是一种滤食性软骨鱼类 , 牠时常被看作姥鲨科成员 , 但是最早的姥鲨属是被发现于南极西摩岛始新世中期地层。Hueneichthys costatus, Rhomphaiodon nicolensis,小弓鲨"Hybodus" minor及哥臣鲨Grozonodon candaui则属于新软骨鱼(neoselachian)最原始的成员并和古鳍棘鲨目Synechodontiformes有密切的关系 , 这些原始的新软骨鱼类和古鳍棘鲨目Synechodontiformes彼此有着相似的齿冠 ,但齿根却是不相似的 ; 至于Reifia minuta及雷蒂亚古鳍棘鲨Synechodus rhaeticus二者的分类地位是存有疑问的 , 二者可能是代表另一个晚三迭纪的软骨鱼类世系 。
于晚三迭纪至早侏罗纪交界时海平面改变并强裂影响西欧 , 瑞替阶(Rhaetian Age)后欧洲中部及北部都被一片浅的陆缘海所覆盖 , 于当时西欧的一片浅海中 , 新软骨鱼(neoselachian)种类占了50% , 牠们于西欧进行辐射形进化 。这时期的西欧存有一些和新软骨鱼(neoselachian)存有关连的短暂软骨鱼类世系例如Vallisia , Doratodus, 伪铠鲨属Pseudodalatias, Raineria 。 伪铠鲨Pseudodalatias是具有切断及压碎食物能力的牙齿 , 这显示牠们能以相对较大的软件动物为食 。
早侏罗纪意大利的Ostenoselache stenosoma有着许多独特的特征包括有较长的脑颅 ,没有背鳍 ,牠的主齿尖和皱鳃鲨属及一些三迭纪的古鳍棘鲨目成员的相似 ; Ostenoselache stenosoma是没有衍生形质(autapomorphic characters) , 对于牠的系统学位置也是不明的。
距马克默多鲨Mcmurdodus后的一亿四千万年后海洋出现了另一类属,古鳍棘鲨类Paleospinax(Synechodus) , 目前对牠们的认识仅来自牙齿化石的发现 , 古棘鲨类Paleospinax是生存于二叠纪早期至始新世时期(约二亿五千万年前至六千万年前) , 但英国及德国仅只发现了小量完整的古鳍棘鲨类颌骨及椎骨化石样本。古鳍棘鲨全长一般小于三呎但和现在的鲨类特征有很多关连 , 例如有较长的吻部,口器已置于头部的下方,短小的颌骨只和颅后相连 , 牙齿有较密的珐琅质及发育良好的椎骨。牠们身体表面存有鳍棘 , 类似现在的角鲨科。尽管对古鳍棘鲨类认知还是很少,但牠们已被认为是属于早期的真正鲨类了。
过去认为古鳍棘鲨目synechodontiform 是于中生代初期出现 , 近来IVANOV 于2005年发表于俄罗斯乌拉尔的二迭纪早期地层发现了古鳍棘鲨属种Synechodus antiquus的牙齿化石 , Synechodus antiquus 的发现显示古鳍棘鲨目synechodontiform于古生代晚期已存在于地球上。
早三迭纪土耳其发现的古鳍棘鲨属种没被定种 , 晚三迭纪诺利克阶加拿大有Synechodus volaticus , Synechodus multinodosus及Synechodus incrementum , 化石都是被发现于英属哥伦比亚省(British Columbia) , 这也显示古鳍棘鲨属于诺利克阶北美旁的大西洋取得分布 。于晚三迭纪雷蒂亚阶(Rhaetian Stage)至古新世达宁阶(Danian)欧洲有广泛的分布 , 第一种出现于西欧是雷蒂亚阶(Rhaetian Stage)法国及英国的雷蒂亚古鳍棘鲨Synechodus rhaeticus 。侏罗纪的属种有早侏罗纪海塔其阶(Hettangian)卢森堡的Synechodus streitzi 及 Synechodus paludinensis ,锡内穆阶(Sinemurian)意大利的Synechodus pinnai , 英国的Synechodus priscus , 侏罗纪中期巴通阶(Bathonian)英国西南部的Synechodus duffini , Synechodus levis , 卡洛维阶(Callovian)玻兰的Synechodus prorogatus , 晚侏罗纪牛津阶(Oxfordian)德国南部的Synechodus riegrafi 及启莫里奇阶(Kimmeridgian)英国的Synechodus plicatus 。
早侏罗纪海塔其阶(Hettangian)至锡内穆阶(Sinemurian) 丹麦,法国及瑞典的Synechodus occultidens , 普连斯巴奇阶(Pliensbachian)英国及瑞典的Synechodus enniskilleni , 托阿尔阶(Toarcian)英国的Synechodus egertonia (Synechodus smithwoodwardi)牙齿形态有别于其它古鳍棘鲨属种而划归新属Palidiplospinax , Palidiplospinax和古鳍棘鲨属的主要不同之处在于其侧齿尖之别。
白垩纪时期古鳍棘鲨属则向北美及亚洲迁移发展 , 白垩纪属种包括豪特里维阶(Hauterivian)至阿尔布阶(Albian)英法的Synechodus dubrisiensis , 阿尔布阶(Albian)至桑托阶(Santonian)俄罗斯及哈萨克的Synechodus dispar , 英国及哈萨克的Synechodus nitidus , 阿尔布阶(Albian)英法及哈萨克的Synechodus tenuis , 坎帕阶(Campanian)美国蒙大纳的Synechodus striatus及加拿大的Synechodus turneri , 于澳大利亚也有古鳍棘鲨未定种的化石记录。
古鳍棘鲨属种于晚白垩纪时期衰落 , 但仍有属种能支持至第三纪 , 比如出现于桑托阶(Santonian)俄罗斯的Synechodus hesbayensis , 牠的化石于比利时晚古新世至早始新世地层都有化石记录, 桑托阶(Santonian)哈萨克的Synechodus lerichei 居然也于比利时古新世地层都有化石发现 , 另一属种Synechodus faxensis于古新世时期丹麦,法国及比利时取得广泛的分布 , 不过牠们于古新世时期仅只局促于西欧 , 于早始新世便完全灭绝了。
原鳍棘鲨属Protospinax是生存于侏罗纪中期至晚期西欧等地的鲨类 , 牠一般全长5呎 , 和鳐类及角鲨目关系密切 , 属种包括侏罗纪中期卡洛维阶(Callovian)至晚侏罗纪提通阶(Tithonian)德国及玻兰的Protospinax annectans , 晚侏罗纪启莫里阶(Kimmeridgean)早期英国的Protospinax planus , 侏罗纪中期巴通阶(Bathonian)英国西南部则存有三种包括Protospinax bilobatus , Protospinax carvalhoi及Protospinax magnus , 此外于法国也有原鳍棘鲨属种化石发现。于德国早提通阶地层就发现了完整的原鳍棘鲨属种骨骼化石 , 牙齿长有卵形的齿冠及突出的低齿尖 , 牙根呈二裂的 , 牠们的牙齿是具有压碎食物的能力 , 这看似原鳍棘鲨属Protospinax是倾向以深海底的甲壳类为食 。有关原鳍棘鲨属Protospinax的分类地位是存有争议的 , 有专家认为牠是最原始的角鲨目成员 , 也有专家认为牠是属于鳐类。最古老的角鲨目成员角锯尾鲨属种Squalogaleus lochensteinensis及Squalogaleus woodwardi也曾被划作原鳍棘鲨属种 , 但后来则被划归角锯尾鲨属Squalogaleus 。
楔齿鲨属Sphenodus (Orthacodus)于侏罗纪的欧洲很常见 , 于欧洲常可找到楔齿鲨属Sphenodus的主齿尖及椎骨残骸化石。楔齿鲨属Sphenodus的牙齿较适合把猎物撕成碎块 , 前牙狭长 , 有较高的尖峰并长有明显的锯边 , 颌前端牙齿外形成S形 , 这些牙齿是处于吻边的并成短且直的折叠。牙根基部平坦 , 并没有任何沟槽 , 牙根和数目众多的不规则血管贯通 。
早侏罗纪属种有欧洲的Sphenodus alpinus , 瑞士的Sphenodus helviticus , 晚侏罗纪属种包括欧洲的提通楔齿鲨Sphenodus tithunius , Sphenodus venulosu , 俄罗斯的Sphenodus stschurowskii , 德国南部的长齿楔齿鲨Sphenodus longidens , 梅加楔齿鲨Sphenodus macer 及光齿楔齿鲨Sphenodus nitidus 。晚侏罗纪时期楔齿鲨属种于南北半球都有分布 , 牠们是大洋性鲨类并可能会随季节迁移回游 。
楔齿鲨属种于白垩纪时期衰落 ,早白垩纪时期仅有Sphenodus salandianus及Sphenodus subaudianus , 晚白垩纪的属种包括森诺曼阶(Cenomanian)的Sphenodus planus 及桑托阶(Santonian)的Sphenodus sennessi , 于澳洲昆士兰白垩纪地层 , 安哥拉坎帕阶(Campanian)地层及南极也发现了楔齿鲨属种化石 , 于南极科尼亚克阶(Coniacian)至坎帕阶(Campanian)地层发现的楔齿鲨属种牙齿化石保存不完整 , 但根据牙齿推断全长至少长达5米。楔齿鲨属种于古新世时期完全绝种 , 最后的属种是丹宁阶(Danian)丹麦瑞典的Sphenodus lundgreni 。
于德国南部发现的长齿楔齿鲨Sphenodus longidens , 梅加楔齿鲨Sphenodus macer 及光齿楔齿鲨Sphenodus nitidus 可带来不少关于楔齿鲨的习性等资料 , 三种不同的牙齿 , 显示牠们的进食方式也有所不同。
梅加楔齿鲨Sphenodus macer有完整的化石被发现 , 牠有较大的胸鳍 , 从梅加楔齿鲨Sphenodus macer盾鳞腹侧被沙石磨擦过的痕迹来看 , 梅加楔齿鲨Sphenodus macer是像扁鲨般的底栖性鲨类。化石样本SMNS 80142/44是一条长85厘米的梅加楔齿鲨 , 牠的牙齿只有8.7 mm , 但是于 Schnaitheim 找到的梅加楔齿鲨Sphenodus macer离散牙齿最大的高度值达18 mm , 这显示化石样本SMNS 80142/44中的个体是未完全成熟的个体 , 成年的梅加楔齿鲨Sphenodus macer全长应该可达1.7米。 从牠的牙齿来看主要以抓紧猎物为主的 , 牠可能是静伏海底泥沙中 , 出奇不意地突袭猎物 。
光齿楔齿鲨Sphenodus nitidus的牙齿高度达22.4 mm , 根据牙齿推测光齿楔齿鲨Sphenodus nitidus全长可能超过 2米。光齿楔齿鲨Sphenodus nitidus牙齿也同样长有锯边 , 但是光齿楔齿鲨Sphenodus nitidus和长齿楔齿鲨Sphenodus longidens前齿主齿尖弯曲有所不同 , 这也显示二者的攻击进食猎物的方式也有所不同 , 光齿楔齿鲨Sphenodus nitidus前齿以捕捉抓紧小型猎物如鱼类及头足类 , 然后把猎物整个吞下 , 从光齿楔齿鲨Sphenodus nitidus和长齿楔齿鲨Sphenodus longidens化石所在来看 , 二者都是属于大洋性的鲨类 , 牠们是生活于海洋中层及表层的鲨类 , 这点和梅加楔齿鲨Sphenodus macer是完全相反的 。
长齿楔齿鲨Sphenodus longidens的牙齿高度达44 mm , 长齿楔齿鲨Sphenodus longidens全长可超过 4米 。长齿楔齿鲨Sphenodus longidens牙齿主齿尖长有锯边 , 牠可能像现存的大白鲨一样 , 透过扭动头部把海生爬行类的肉撕扯下来 , 因此长齿楔齿鲨Sphenodus longidens牙齿主齿尖大多折断 , 长齿楔齿鲨Sphenodus longidens更曾被看作是应归属鼠鲨目。长齿楔齿鲨Sphenodus longidens于晚侏罗纪的英国及德国 , 晚白垩纪森诺曼阶(Cenomanian)至桑托阶(Santonian)分布于日本 , 巨大的体型和锯齿切边显示牠可能以小型海生爬行类为食, 恐怕是史上第一种具掠食性的鲨类。
副楔齿鲨属Paraorthacodus于侏罗纪世界各地留下许多牙齿和椎骨化石 , 副楔齿鲨属Paraorthacodus有较大及长长的主齿尖 , 主齿尖及侧齿尖都有锯边 , 牠们的牙齿是以抓捕猎物为主的。副楔齿鲨属从侏罗纪兴起 , 至始新世时期才完全灭绝。
副楔齿鲨属Paraorthacodus的分布和六鳃鲨目成员Notidanodon都很相似 , 尽管白垩纪时期副楔齿鲨属种的种类多样性并不高 , 但却于两极气候和暖及寒冷的地区都有分布 , 晚白垩纪时期副楔齿鲨属种于南极都有分布。从副楔齿鲨属Paraorthacodus的化石形态来看牠们和须鲨目同样生活于浅海的鲨类 , 以鱼类及软体动物为食的缓慢鲨类。
副楔齿鲨属种包括晚侏罗纪德国的Paraorthacodus kruckowi , 启莫里阶(Kimmeridgean)至提通阶(Tithonian)的Paraorthacodus jurensis , 白垩纪 阿尔布阶(Albian)英国,法国及俄国的Paraorthacodus recurvus , 坎帕阶(Campanian)蒙大纳的Paraorthacodus andersoni , 桑托阶(Santonian)至坎帕阶(Campanian)哈萨克,德国及比利时的Paraorthacodus nerviensis , 晩白垩纪阿根廷的Paraorthacodus patagonicus ,晩白垩纪桑托阶(Santonian)至坎帕阶(Campanian)南极北部的南极副楔齿鲨Paraorthacodus antarcticus , 麦斯特里希特阶(Maastrichtian)瑞典的Paraorthacodus conicus , 晚白垩纪至古新世新西兰的Paraorthacodus sulcatus及Paraorthacodus validus , 晚古新世至始新世北美及英国的Paraorthacodus clarkii及始新副楔齿鲨Paraorthacodus eocaenus则为最后的副楔齿鲨属种
在本文为化石网论坛精华帖,由网友景德泰岚提供,仅供大家学习参考。
编辑:刘琮滢