海洋起源与演化 origin and evolution of the oceans
海洋起源与地球起源密切相关。近两个世纪以来,人类有关海洋起源与演化问题的知识已取得很大进展。尽管问题还未得到解决,然而却有可能根据现有的资料(主要是地质记录)和现代的地球史观对海洋的发展提出一些模式。
一般认为地球远在46亿年前起源于原始太阳星云,它是通过吸积星云物质(一些气体和尘埃微粒)和经碰撞等物理演化过程而形成的。地球在形成时,温度较低,几乎是均质的。吸积造成了地球的致密与收缩,而收缩则使原始星云物质相互摩擦,从而释放出一定的热能;放射性物质的早期衰变释放大量的热能;陨石类物质撞击地球也能造成局部性的升温。这些作用使地球普遍升温,造成了熔融,地球便在此全球性的热事件中,并在重力作用之下,分化为地核、地幔和地壳的层状结构。这一过程既决定着岩石圈的演化历史,也影响着大气圈、水圈以及生物圈的生成与演化。
海水起源与演化 在地球历史的早期,大气圈与水圈密不可分,当时的水分呈气态(蒸气)混于原始大气之中,所以有关海水早期历史的推论也必然集中于早期大气圈历史的探讨上。
大洋水的形成 在地球演化早期,由星云凝聚而成的原始地球一方面继续吸积、扩大,另一方面气体与固体粒子逐渐分开,形成原始的大气圈。但原始大气不断向星际空间逸散,所以原始大气圈对于地球水圈的形成并无贡献。在原始大气逸散的过程中,来自地球内部的挥发物质不断替补着原始的大气,逐渐演变成一个次生的大气圈。水圈的形成起初便有赖于这个次生的大气圈。次生大气圈的发育主要来自象火山喷发这样的释气作用。当时它是还原性质的,而不是氧化性质的(氧化是现代大气圈的特征)。次生大气圈生成之初,地球温度还较高,几乎所有的挥发气体,包括 H2O都在大气之中。但当地表温度降到沸点以下,H2O便冷凝为水,形成原始海洋。由于大气中含有HCl和CO2等气体,故早期冷凝出来的水必定是酸性的,温度也较高。
原始大洋是酸性的热洋。原始大洋的酸性热水与火成岩中的矿物发生剧烈作用,溶解出二氧化硅和某些阳离子,造成主要是由含铝粘土矿物所组成的沉淀物,这便是原始大洋的沉积物。导致原始大洋发展的化学过程可归结为:原生火成岩矿物+酸性挥发物+水→沉积岩+大洋+大气。
根据地球上已发现的古老沉积岩年龄,可推测地球水圈的出现可远溯至地质历史早期。在南非发现了年龄为34亿年的古老沉积岩,证明当时地球上已有水存在;该岩石中还发现极原始的有机体遗迹,说明水的出现可能更早。现今已发现的最古老岩石位于格陵兰西南部,年龄约38亿年,岩石虽经变质,仍然保留了沉积作用的痕迹,提示当时水已出现。因此推测,地球水圈形成于35~38亿年前。
大洋水是漫长地质历史中地球内部排气的产物。随着时间的推移,大洋水因排气作用不断积累增长。据不同学者推测,水量增长有 3种可能的方式:①早期增长较快,在水量-时间坐标图上显示为抛物线型,即认为大部分海水是在地质历史早期形成。②匀速增长的直线型。③晚期增长较快的指数曲线型,即认为大部分海水是在地质时期后期出现的。
早期大洋水 地球水圈形成后,距今35到20亿年,也许延续到距今15亿年,是酸性、缺氧的原始大洋水逐步向现代大洋水过渡的时期。当时大气中的含氧量大大低于现在,大洋水中的氧分则更低,沉积岩的来源也似乎比后来的岩源更为基性。矿物、岩石在缺氧大气中崩解,变为碎屑,岩屑又在主要是缺氧的海洋条件下堆积起来。缺氧的证据主要有:①20亿年以前的碳酸盐是很少见的,这个时代的碳质沉积主要以石墨和油母岩的形式出现,证明当时缺少游离氧;②18~20亿年以前的沉积岩中有许多易氧化矿物的碎屑,如黄铁矿和沥青铀矿的碎屑等。这些碎屑虽然具有磨损、滚圆等浅水拖动的特征,但是它们并没有遭到氧化;③距今32~20亿年的古老地层中广泛出现沉积韵律清晰的条带状铁矿,它们是从水体中沉淀出来的化学沉积物。铁的大规模搬运只能在缺氧环境下以亚铁形式(还原状态)进行。但当时生命已在海洋中出现。厌氧生物的光合作用产生了少量生物性氧,它把溶解在海水中的亚铁氧化为正铁而沉淀下来,韵律性的条带显示了生物性氧的数量变动。总之,早期大洋水是缺氧的,而且可以把它看作是玄武岩经酸性淋滤而产生的溶液。
现代大洋水 早期大洋水的pH值较现代海水为低。那时的火成岩偏于基性,当它们与酸,如HCl作用时,会产生中性或偏碱性的溶液,原始的酸性大洋便逐步改变其性质。早期大洋水中的负离子可能主要是碳酸根和碳酸氢根离子,而不是氯离子,但随着 HCl对岩石的淋滤作用,造成硅的释放与沉淀,并不断产生氯化物,早期大洋便逐渐向含氯离子的现代大洋过渡。大洋水可能在距今15~20亿年时开始具有它现代的特点。这时海洋中具真核细胞的绿色植物出现,生物造氧能力大大增强,大量游离氧生成,氧不仅可以满足海洋中的各种氧化反应,而且可以从水中外逸至大气中,逐步形成具有现代特点的大洋水和大气,以及高度氧化的地面条件。游离氧的出现使原来呈亚铁溶液状态的大规模搬运和生成条带状铁建造的沉积过程基本停止。铁被氧化为正铁,不再溶于水,只能保存在风化剖面中,形成15~20亿年以来地球上广泛出现的红层。这是水圈和大气圈开始具备现代特点,充分氧化的重要证据。从这时起,有足够的氧能使二氧化碳(CO2)转化为碳酸根(CO-23),使还原性硫(S)转化为硫酸根(SO-43),因而碳酸盐岩(主要是碳酸钙──灰岩)和部分硫酸盐岩(主要是硫酸钙──石膏)开始大规模形成。距今20~15亿年以来,各时期所形成的各种沉积岩类型之间的数量比率大致是相同的。
长时期以来,人们简单地认为现代大洋的盐分是来自于岩石的盐分的积累,岩石被风化,经河流搬运,使盐分不断汇入大洋,因而大洋水的盐度应该随着时间的推移愈来愈高。然而,大洋水体并非是盐的简单积聚场所。在水和某些盐类向大气圈蒸发的同时,某些盐类也必然以形成矿物的方式从水体中析出,转移到岩石圈中;或是被生物吸收,转移出水圈。所以现代大洋在化学总体上长期维持着一种盐类收支平衡的稳定状态,它不是盐类的单纯累积区,而是大陆(和很少部分的大气)物质向海底传输和部分再返大气的中间站。大洋成为稳态系统的重要特性早在15~20亿年前便已开始形成。

洋盆的起源与演化 大洋盆地是海水汇聚的地方,它的形成与发展是海洋起源与演化的重要方面。
洋盆起源 历史上曾出现过各种各样有关洋盆起源的学说。陨石撞击说认为,大洋是由于巨大的陨石撞击地球产生一次或多次爆炸而形成的;月球分离说认为,月球是从地球母体分离出去的产物,而月球从地球上撕开时所留下的巨大伤疤,便是现代的太平洋;地壳均衡说认为,地壳密度小的区域形成大陆,密度大的形成洋盆;海底扩张与板块构造说为洋盆的起源与演化问题,提供了比较合理的解释。海底扩张说认为,地幔岩浆在大洋中脊处上升并固结,生成大洋地壳,较老的地壳被推向两侧从而生长出洋盆。新洋壳不断生长,其生长率为洋脊两侧扩张速率之和。宽达 1.5万公里的太平洋洋底只需 1.5亿年左右即可形成。这与观测到的洋底年龄相符。新洋壳的不断生成必须同时在另外的地方伴随有老洋壳的破坏与消亡,否则地球将无限膨胀起来。深海沟是较老的、逐渐冷却的洋壳向下俯冲、返回地幔的地方。板块的分离和汇聚造成大洋发生、成长与消亡的历史。
洋盆的发展阶段 J.T.威尔逊等将大洋盆的演化划分成两大阶段,即成长阶段与衰退阶段。每个阶段又各分三个时期:成长阶段分为胚胎期、幼年期和壮年期,衰退阶段分为衰老期、终结期和残痕期(见表)。有人将上述演化阶段称为威尔逊旋回。
胚胎期的典型代表是东非裂谷系,它具有许多与大洋中脊类似的特点,虽然它还没有生成洋壳,但已张裂,可视为胚胎型的大洋。幼年期的典型代表是红海、亚丁湾,为狭窄的年青洋盆,已出现新生洋壳,是现代大洋的雏型(见彩图);挪威海、巴芬湾也是幼年期的实例。壮年期的典型代表是大西洋,它约在侏罗、白垩纪开始张开,发育至今已十分成熟,具有现大洋的一切特点,中部为大西洋中脊,中脊轴部有裂谷,裂谷内不断溢出枕状、绳状拉斑玄武岩;印度洋与南大洋也是壮年期的例证。衰老期的典型代表是太平洋,它被视为泛古洋(panthalassa)的残余,在二叠纪和三叠纪时期,泛古洋覆盖着泛古陆(pangaea)以外的整个地球表面。太平洋是世界上最大的大洋,它的扩张速率很高,但因洋盆边缘收缩的速率大于东太平洋海隆的扩张速率,使它正处于总体收缩的过程中。太平洋边缘发育着一系列岛弧、海沟和年青的褶皱山脉,周围大陆不断向洋盆掩覆而来,使之缩小。终结期的代表是地中海,这个海盆在中生代时期就已存在。由于非洲与欧亚板块的相对运动,至少在中新世时就曾被封闭过,地中海干枯,变成内陆沙漠,形成大量的岩盐等蒸发岩;距今约500万年时,直布罗陀海峡被冲破,大西洋水复又灌入地中海。黑海和里海也是这类例子。残痕期的代表是喜马拉雅山,而印度河-雅鲁藏布江构造线,便是大洋封闭后留下的疤痕──地缝合线。在中生代时期,这里曾经是分隔冈瓦纳古陆(Gondwana-land)与劳亚古陆 (Laurasia)的特提斯海(Tethys)的一部分。
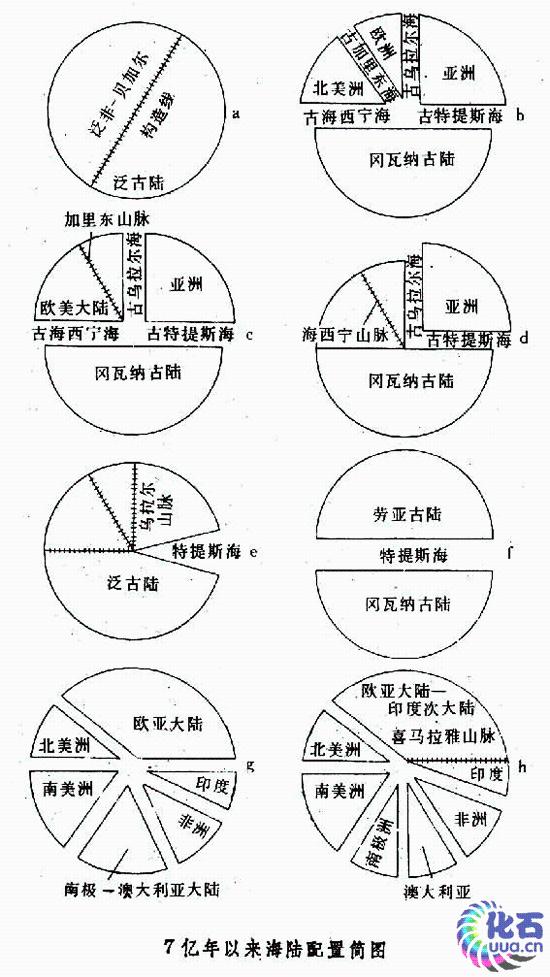
洋盆演化史 在地球演化的历史中,大洋与大陆经常改变它们的地理位置、配置形式,以及它们的大小与数量。洋底扩张、大陆漂移,即板块分裂与聚合的作用至少已有 7亿年了,它们也许还可能有长达20多亿年的历史。在过去的 7亿年历史中,大洋与大陆的相对轮廓、配置形式和变化状况留下了比较明显的痕迹,可供人们对它作初步的探索。大约7亿年前的元古代末期,地球上只存在着一个超级大洋──泛古洋和一个超级大陆──泛古陆,泛古陆是由以前分离的大陆组成的,泛非-贝加尔构造线便是它们的板块缝合线(图a)。距今 5.7亿年的寒武纪,泛古洋继续存在,它可能就是太平洋的前身。当时地球上存在着 4个大海和 4个大陆,它们是古海西宁海、古加里东海、古乌拉尔海和古特提斯海,北美洲、欧洲、亚洲和冈瓦纳古陆(图b)。距今 3.7亿年左右的泥盆纪,古老的欧洲与北美洲板块碰撞,合并成欧美大陆,古加里东海消亡,出现加里东山脉(图c)。距今 3亿年的晚石炭纪,欧美大陆沿海西宁构造带与冈瓦纳古陆连接起来,古海西宁海消亡(图d)约2.25亿年前的二叠纪,亚洲沿乌拉尔构造带与欧洲大陆合并,重新组成泛古陆,古乌拉尔海消亡。但古特提斯海仍然存在,它改变形态,以一个三角形的海楔入泛古陆之中,再逐步演化成后来的特提斯海(图e)。这时作为太平洋前身的泛古洋仍继续存在。距今大约1.9亿年的中生代──侏罗纪早期,从墨西哥湾到直布罗陀一线张裂开来,承袭泛古洋的太平洋分出一支切入墨西哥湾,称为太平洋海。它与由东向西而发育伸展过来的特提斯海相连在一起,把一个超级大陆──泛古陆分隔成两个大陆,即北边的劳亚古陆和南边的冈瓦纳古陆(图f);这时太平洋已正式形成,现代太平洋西北部洋底所发现的年龄近 2亿年的洋壳就是这种证明。距今 1.35~1.1亿年间的早白垩世,南大西洋开始张裂,生成洋盆,而后欧洲与北美洲分裂,因而冈瓦纳古陆和劳亚古陆也开始破裂。在距今 0.7亿年的晚白垩世,劳亚古陆和冈瓦纳古陆都高度分裂,大西洋、印度洋和南大洋相继出现;两个古陆崩解后形成了欧亚大陆、北美大陆、南美大陆、非洲大陆、印度次大陆和南极洲-澳大利亚(图g)。从距今0.65亿年的古新世以来,格陵兰与斯堪的纳维亚分裂,挪威海与格陵兰海形成,北冰洋与大西洋连通。距今0.53亿年的始新世初,澳大利亚裂离南极洲,并开始向北漂移;距今 0.4亿年的始新世晚期印度与亚洲主体碰撞,特提斯海东段关闭(图h);约0.18亿年前的中新世期间,阿拉伯与亚洲主体碰撞,特提斯海西段部分关闭,遗留下西通大西洋的地中海;约 0.035亿年前的上新世时,北美与南美重新相逢连通,巴拿马地峡升起,切断了大西洋与太平洋的直接通道,强烈地改变了大洋水的环流系统。
对于7亿年以前海洋演化史的认识是非常模糊的,这是由于人们对地球早期的历史缺乏认识,有关前寒武纪地质发展史的争论还很大。在人们尚未确切探明地球的起源以前,关于海洋起源的任何解释和学说都带有很大的推测性。
生命与海洋 生命孕育于海洋之中,生命的起源和早期演化依赖于大洋的发生和演化。大洋对于生命的形成提供了必要的物质基础和生存环境。
尽管一切有关生命起源的讨论是推断的,但是生命在地球历史上开始得很早,并发生于大洋之中则是有证据的。年龄约34亿年的古老海洋沉积岩中发现了细菌化石遗迹,故生命的开始必定在此之前。生命的起源应始于氨基酸的形成,而最早出现的氨基酸却是无机合成的有机化合物。实际上,作为生命前驱的一些分子都是无机合成的,它们当中有组成蛋白质的氨基酸,组成核酸的糖和碱基,以及构成膜的脂类等分子。早期还原性大气圈的一些气体溶解于大洋水中,使原始的大洋好似一种稀汤,在放电和紫外线辐射等能量作用之下,它们聚合成氨基酸、碱基和核糖等大的有机分子,目前在实验室里已证实了这种合成方式。这些合成的有机单体在热力学上有利于解聚作用的大洋中再次聚合,最后便组成蛋白质、核酸等生命物质。虽然紫外线的辐射能量促使了生命物质的合成,但是在早期缺氧的大气中并没有足够的游离氧去形成能吸收紫外线的臭氧隔离层,强烈的紫外线对于在地表刚形成的生命物质具有巨大的杀伤力,因而初生的生命物质必须依靠大洋,生存在紫外线达不到的大洋水中。对于初生的生命物质和原始生命,氧是一种具有侵蚀力的毒性气体。一旦出现游离氧,氧化作用就将破坏氨基酸等生命物质,破坏如厌氧细菌等没有保护系统的原始生命体,故而一个还原性的环境乃是从无生命的有机物进化到生命物质和原始生命体所绝对必需的条件。早期缺氧的大气和大洋正好提供了这个条件,生命物质便得以在大洋中积累起来,演化成生命。所以大洋是生命的摇篮。
大洋中,也是地球上,第一批出现的生物是些单细胞的实体,它已初步具有遗传的机构。前寒武纪大洋中生命演化的第一个重大事件是光合体的出现,最初的光合生物,出现于距今约30亿年以前。光合体合成有机食物维持自己的生存,但又释放出能破坏自身的氧。它产生的氧大部分被当时大洋中的亚铁溶液所吸收,形成大规模的条带状铁矿建造,但是在光合体形成以前已经出现了可以抵御氧对生命体侵蚀的间氧酶。这个时期的生物是原核细胞组成的,即原核生物,它们只能无性繁殖,主要是细菌和蓝藻。前寒武纪生命演化进程中的第二个重大事件是有核细胞的形成,约发生于18~14亿年前。有核细胞组成的生物,即真核生物,具备有性繁殖能力。有性繁殖使新获得的适应性通过群体迅速传播开来,因而大大加速了生物进化与变迁的速率。
距今15~20亿年前,地球大气圈和水圈的性质经受了质的变化,从很少或没有游离氧的还原性大气与大洋,变成了氧化的大气与大洋。臭氧层的形成大大减少了地球表面的紫外辐射,生命的演化无需再隐蔽在大洋的深处进行,它们可以逐渐转移到浅水处,甚至陆上。而光合体和有核多细胞的相继出现,使新的生命体不再惧氧,它们可以在富氧的环境中直接依靠可见光的能量进行光合作用,产生更多的有机物。10亿年前,真核生物已是十分繁多,而5.7亿年前开始的寒武纪更是急剧地涌现出大量种类繁多而又复杂的有壳多细胞型生物。从此,生物便真正开始其丰富多彩的进化阶段。