三趾马动物群化石
三趾马动物群是已知种类包括啮齿类的原鼬鼠、副竹鼠、甘肃豪猪,食肉类的短吻犬、中华副美洲獾、近狼獾、小原臭鼬、霍氏原臭鼬、大密齿獾、中华貂、鼬鬣狗、似鬣狗鬣形兽、翁氏鬣形兽、变异副鬣狗、巨鬣狗、巴氏剑齿虎、野猫、小后猫,长鼻类的保德四棱齿象,奇蹄类的东乡三趾马、贾氏三趾马、渭河三趾马、腔脊三趾马、膜鼻三趾马、和政无鼻角犀、维氏大唇犀、伊朗犀、简单副板齿犀、临夏副板齿犀、林氏额鼻角犀、双角犀、钩爪兽,偶蹄类的斯氏弓颌猪、弱獠猪、叉角鹿、后麂、新罗斯祖鹿、萨摩麟、舒氏河南兽、小齿古麟、羚羊、步氏和政羊、中新羚、中华羚、原大羚、陕西旋角羊等,还发了临夏鸵鸟化石。
临夏鸵鸟:临夏鸵鸟一种巨型鸟类,比现在的非洲鸵鸟还大,鸵鸟是现存鸟类中体型最大的鸟类,它的前肢已经十分退化,胸骨不具龙骨突,没有飞行能力,两趾行走。甘肃鸵鸟的材料代表了早期鸵鸟保存最好的骨骼标本之一,这一发现也将鸵鸟在中国生活的历史向前推移数百万年。在鸵鸟化石中,临夏鸵鸟具有迄今为止保存最好的腰带。临夏鸵鸟的发现表明晚中新世鸵鸟已经广泛分布于非洲和欧亚大陆。新的鸵鸟化石的发现不仅对该地区的气候环境具有一定指示意义,而且对探讨鸵鸟类的早期演化提供重要价值。
原臭鼬:原臭鼬是一种在欧亚大陆广泛分布的新近纪鼬科动物,但以前建立的种都是基于一些单个牙齿。在和政地区发现的大量完整的头骨以及骨架化石,使古生物学家对于原臭鼬的认识在质量、数量和种的多样性上都前进了一大步,并为解决原臭鼬的系统发育和地理分布难题提供了绝好的机会。小原臭鼬是2004年在和政地区发现的一个新种,是世界上最小的原臭鼬,它的牙齿和骨头构造也最原始。原臭鼬适应于欧亚大陆中纬度地区的开阔草原环境,它在晚中新世时期的中国具有与欧洲相同的多样性。

原臭鼬
印度熊:印度熊具有巨大的头骨和粗壮的牙齿,裂齿已经失去了切割的功能,臼齿变成方形,上面有钝的齿尖。和政地区的印度熊标本相当丰富,而且保存得非常完好。熊类的尾巴退化到仅有残迹,腿脚变得短而笨重,因此它们追捕猎物的能力明显减退。现代的熊更是变成了杂食性动物,与狗类以肉为食的习惯相差甚远。熊类在北半球起源和演化,在更新世时期侵入南美洲,但它们却奇怪地从未侵入过非洲。熊类有各种各样的生活环境,如冰天雪地中的北极熊和晚更新世时住在山洞中的洞熊。
副鬣狗:副鬣狗的体型与现代的鬣狗接近。鬣狗其貌不扬,其后腿很短,跑起来有些颠跛。鬣狗常给人一种贪婪的印象,它们成群结伙抢劫其他动物的猎物,有着吃腐肉的恶名。其实现代鬣狗是奔跑追击型捕猎动物,以穷追并群体合作的方式追捕猎物,它们的食物比例中以自己捕食所得的新鲜猎物为主,抢占别人的猎物只占很小部。鬣狗与其他食肉类最大的不同之处在于其特有的粗壮上下颌骨及前臼齿,可轻易咬碎骨骼,从中摄骨髓。一群鬣狗可以在几分钟内吃光一整只斑马,连骨带肉丁点不剩。
鼬鬣狗:鬣狗的四肢因经常奔跑而变长,牙齿和颌骨因咀嚼骨头而增大。牙齿的增大尤其表现在最后两枚前臼齿,也就是第三和第四枚锥形的前臼齿上,这是鬣狗咬碎大骨头的很需要工具,因为它们在习惯的食腐肉的过程必须对付那些难啃的骨头。鬣狗虽然看起来有些象狗或狼,但它们不是真正的狗,与狗的亲缘关系比较远,倒是与猫的关系更近。鼬鬣狗是早期原始的鬣狗,比后期的鬣狗小而轻巧,是晚中新世和早上新世常见的一种小型鬣狗,在华北各地这一时期的动物群中发现过很多它们的化石。
鼬科:鼬类的特点与其它的狗形类相差较远,它们从渐新世初期起源后就沿着一条独立的道路发展。鼬类的脸部短,脑颅长而扩大,这是它们的典型特征。从其祖先类型开始,鼬类新生代中、晚期在发展上出现了高度的适应辐射,其中有不少生存时间很短就灭绝了的类型。直到今天,还有鼬类的五个类群生活在世界上,分别以黄鼠狼、蜜獾、獾、臭鼬和水獭为代表。鼬类在所有的食肉动物中表现出了最大范围的适应辐射。和政地区的三趾马动物群中丰富的鼬类动物,包括副美洲獾、近狼獾、原臭鼬、密齿獾和中华貂等。
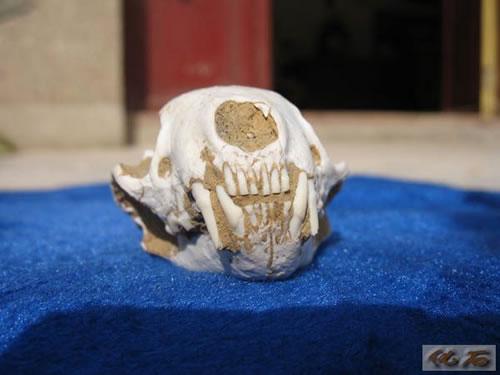
臭鼬
剑齿虎:在凶猛的猫科动物中,剑齿虎是中型或大型的非常特化的类型,剑齿虎的大小与现代的狮子或豹子差不多。作为猫科动物的一员,剑齿虎与它的现代亲戚一样,具有可收缩的锋利的爪子,其牙齿数目减少,上下裂齿的刃叶长而且非常锋利。剑齿虎最大的特点是其长而侧扁弯曲的上犬齿,形同匕首一般,前后还具有锋利的刃嵴,这就是其名称“剑齿”的来源。相反,剑齿虎的下犬却退化缩小了。剑齿虎下颌骨的最大特点是具有非常明显的颏突,通俗地说,就是它的下巴颏特别发达,几乎成一个直角型。



剑齿虎
四棱齿象:长鼻类在它们的全部历史中都是森林中或平原上的大型或巨大的哺乳动物。和政地区有丰富的长鼻类化石,尤其是在中中新世。到晚中新世时期,随着环境的改变,长鼻类在和政的数量减少,仅以四棱齿象为代表。四棱齿象的下颌也具有由门齿形成的铲板,比铲齿象的铲板更窄。中国古代就开始采集来作为中药的“龙骨”中就包括了四棱齿象的化石。长鼻类沿着不同的方向演化,它们有着复杂的进化历史,许多种类最终灭绝了,现在仅仅剩下亚洲象和非洲象还残留在世界上。
三趾马:三趾马具有高冠和白垩质覆盖的颊齿,随着高冠齿的磨蚀,釉质的复杂结构就暴露在咀嚼面上。尽管这些釉质结构在年龄和个体之间都有变化,但不同的属种也具有自己独特的样式,可以作为鉴定的重要标志。三趾马的上颊齿原尖的内侧,也就是靠近舌头的一侧。与现代马相比,三趾马的四肢更为细长。最早的中趾马在120万年前的中新世晚期开始时第一次跨越了白令陆桥从北美来到亚洲,然后迅速扩散到欧洲和非洲。
林氏山西犀:山西犀在分类上属于无角犀亚科,但它却是鼻子末端长着一个小角的犀牛。山西犀最早的材料是由19世纪未来中国的德国传教士从天津的中药材铺买到的,是一些单个牙齿。在和政地区发现了山西犀的完整头骨化石,产于距今500多万年前的早上新世红粘土堆积。山西犀是从无鼻角犀向大唇犀演化的一个中间环节。山西犀也像大唇犀一样有一个扩展的下颌联合部和獠牙状的下门齿,但扩展的程度和獠牙的尺寸都明显小于大唇犀。山西犀的高齿冠和细密的釉质褶皱还显示它是一种以硬草为食的犀牛。
伊朗犀:伊朗犀是一种大型的犀牛,正像其名字暗示的那样,它第一次被发现于伊朗。它的鼻骨又宽又长,用以支撑巨大的鼻角,雄性个体的鼻角尤其发达。它的眼眶强烈向外突出,其目光在生活时一定显得非常威猛。伊朗犀是迄今为止唯一发现在雄性个体的颧弓后部具有粗糙的半球形结构的犀牛,显示其可能附着类似于“角”状的皮肤衍生物,是雄性在与捕食者对抗或与同类争斗时的武器或恐吓工具。伊朗犀雄雌个体之间具有显著差异,即“性双形”现象,说明这种犀牛在北半球开阔的温带草原上过着一雄多雌的生活。
临夏副板齿犀:副板齿犀是一种大型的板齿犀,与进步的板齿犀不同,副板齿犀颊齿的釉质褶皱很弱。副板齿犀到目前为止只发现于中国,但以前发现的材料都相当少,仅是一些单个的牙齿和肢骨,而在和政地区发现的副板齿犀则有相当完好的头骨材料。副板齿犀的头骨相当长,颊齿列强烈后移以至于最后一枚牙齿也位于眼眶之前。鼻骨的游离部也相当长,并形成穹隆状的粗糙角座,显示它在生活时有一个巨大的鼻角。与第四纪的真板齿犀不同,副板齿犀没有额角,真板齿犀具有一个高耸的额角,却没有鼻角。

临夏副板齿犀
爪兽:爪兽是一种很奇怪的奇蹄动物,外形很像马,牙齿与雷兽相似,最特别之处在于它是有蹄类动物中唯一的长爪子的类型。爪兽身高可达3米,常用指关节行走,以保护长长的爪子,它那强有力的四肢是非常有效的防卫武器。但是更多的时候,爪兽用它的爪子勾下树枝,以便吃到最鲜嫩的树叶。爪兽的身体比例看起来相当奇怪,跟大猩猩很相似,其前后肢之间在长度上的差别很大,后肢很短,说明其奔跑能力很弱。从爪兽的肢骨所反映出的特征表明爪兽以适合于承重的后肢作支撑,用灵活的前肢抓取树木的枝叶来食用。
小旱獭:小旱獭是一种个体很小的旱獭,它是在和政地区发现的一个新种。我国的现生旱獭分布于甘肃、青海、新疆、四川、云南和西藏等地区,它们均栖息于高山草原或草甸地带。例如,喜马拉雅旱獭主要栖息于海拔3750~5200米之间的高山草原、高山草甸、谷地灌丛草原、高原荒漠草原、高原高寒荒漠及山地荒漠等环境中。小旱獭在形态上和现在旱獭形态上和现生旱獭非常接近,它们的生态环境也应该大体相同,表明和政地区在早更新世可能已有适于旱獭生活的高山草原或草甸的生态环境。

小旱獭
安氏猴:灵长目是包括我们人类在内的一个分类单位,之所以叫灵长目,是因为人类看作是进化历史的最高产物的自然结果,即是世界上的万灵之长。然而,从进化的过程来说,灵长类是直接从原始的食虫类祖先起源的,所以在整个哺乳动物进化树上,灵长类是想当早的一个分支。中国西北地区发现的灵长类动物非常长,而和政地区却在早更新世的真马动物群中发现了猕猴和副长吻猴的化石,这是非常难能可贵的。安氏猴的个体比现生猕猴各种和化石种稍大,它是我国最早建立的猕猴属的一个种。

甘肃副长吻猴:甘肃副长吻猴是在和政地区发现的一种新的灵长类动物,以产地甘肃为其命名。它的牙齿在尺寸上比所有已知猕猴各种都大,而与原黄狒接近。原黄狒在头骨和下颌上与猕猴很接近,但在牙齿上却与狒狒接近,可能是这两个属共同的祖先。副长吻猴的主要分布区是欧洲,其尺寸大,头长超过20厘米中,面部长,与猪猡接近,雄性犬齿特别发育。和政的材料显示副长吻猴和原黄狒确实可以区分开来,副长吻猴的个体比原黄狒大。甘肃副长吻猴是一种向摄食比较干硬食物方向特别的副长吻猴。
德氏犬:在龙担发现了大量犬属的化石,包括雄性和雌性个体的几十件标本。德氏犬是根据龙担的材料建立的一个新种。十分奇特的是,在龙担动物群中还生少着另外两个犬属的新种,即龙担犬和短头犬,但个体数量较少。德氏犬种名献给法国神父德日进,他不但描述过我国众多地点的犬属化石,而且预见到我国早更新世地层中可能还有另一种没有命名的犬类动物。德氏犬是一种大型犬,雌性的头基长平均19.5厘米左右,雄性长21厘米左右。龙担犬和短头犬的个体也是相当大的。
德氏狗獾:德氏狗獾的个体比大部分现生狗獾稍大,头基长约为12厘米。欧亚大陆的狗獾化石非常少,在南亚和中亚就没有它们的记录,而在欧洲材料比较好的化石实际上只有两种,都与中国的狗獾不同。有趣的是,德日进在1945年讨论贾氏狗獾时就指出过在中国的早更新世应该有另外一种狗獾,但德日进当时并没有建立新种,主要是因为他的材料大部分都是下颌,头骨和上牙很少,龙担发现的完好材料证实了德日进的预见,为了纪念他,所以将在龙担发现的狗獾新种命名为德氏狗獾。
锯齿虎:锯齿虎的分布非常广泛,在亚洲生活于上新世到中更新世,在欧洲生活于上晚新世到中更新世,在北美生活于晚上新世到晚更新世,在非洲生活于晚上新世。锯齿虎是中至大型具带锯齿马刀形上犬齿的猫类动物,它的门齿、犬齿、上第四前臼齿及下第四前臼齿和第一臼齿的齿缘未磨蚀时有锯齿,四肢骨比较细长。锯齿虎作为属名首先是在1890年提出来的,所依据的标本是产自意大利的一个头骨的前半部和下颌,这件标本现在保存在佛罗伦萨大学的地质古生物研究所。
泥河湾巨颏虎:巨颏虎在世界上的分布相当广泛,在亚洲、欧洲、非洲和北美的上新世或更新世地层都有发现。在和政地区发现的泥河湾巨颏虎是中等大小具微弯匕首状上犬齿的猫类动物,其牙齿均无锯齿,四肢骨比较粗短,但比美洲剑齿虎者细长。在中国最开始是德日进于1930年在河北泥河湾发现的,他定了一个新种泥河湾巨颏虎,作为这个种选型标本的一个比较完整的头骨就保存在天津自然博物馆。欧洲的巨颏虎的头骨材料很零散,意大利正型地点材料也很破碎,没有完整的头骨。
临夏西瓦猎豹:西瓦猎豹是中等大小的猫科动物,四肢骨修长,体型大于现生猎豹。在和政地区龙担发现的材料使研究者更加相信西瓦猎豹确实和现生猎豹有很显著差异,例如所有的归入西瓦猎豹的化石都明显比现生猎豹大。临峡西瓦猎豹以化石产地所在的临夏盆地命名,它是大型的西瓦猎豹。西瓦猎豹过去发现的材料主要是下颌,头骨材料很少。此前仅在法国的一个地点发现过这类动物的头骨。我国早期的猎豹化石过去只发现过下颌,和政地区发现的完整材料极大地丰富了古生物学家对这种猫科动物的认识。

临夏西瓦猎豹
龙担日本鹿:日本鹿是中等大小的六角枝鹿类,即它的每一个角是有三个角枝,其眉枝高,主枝长而直。日本鹿的角细长,断面圆或稍扁,角面纹饰以沟为主,第二分叉非常高,第二、三角枝在长短和粗细上相差不大。龙担日本鹿以和政地区的化石地点龙担命名,是一种大型的日本鹿。龙担日本鹿眉枝这一点足以使它和现生的每角三角枝的水鹿和轴鹿区别开来。日本鹿的眉枝与主枝间的夹角大,且较弯曲是与水鹿进一步区别的特征。与所有已知日本鹿的各种相比,龙担日本鹿都是最大的。
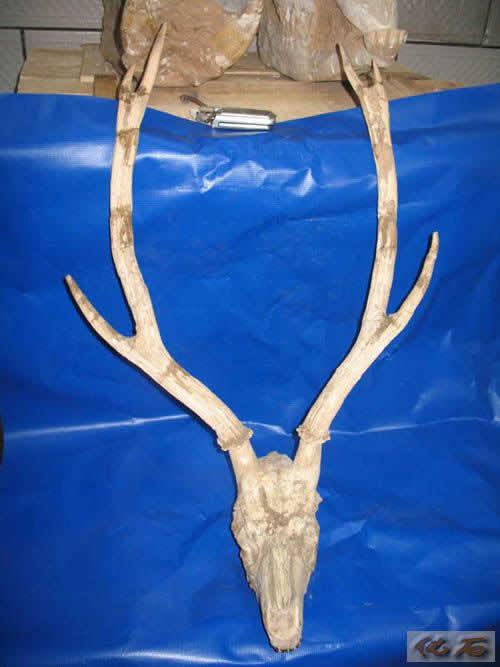
龙担日本鹿
短角丽牛:丽牛是一种头骨低、窄,四肢较细长的牛类动物,在更新世早期分布于欧亚大陆。丽牛的雌性无角,雄性的角较细,亚柱状,角基起始于眼眶稍后,角基至眼眶后缘之间的距离和眼眶本身长大约相等,角的下缘在枕骨顶面之下,角向后伸,不同程度地互相分离,丽牛的枕面近半圆形,头骨在角后的部分仍然很长。在和政地区发现的短角丽牛体型大,鼻骨和额部宽阔,其雄性具一对短而较直或微向内弯、主要向后平伸的角,角的断面呈横长的椭圆形。

短角丽牛
秀丽半牛:半牛是一种头骨较宽的牛类动物,在更新世早期分布于亚洲地区,半牛的角长,角内没有大的空腔,其断面呈三角形,两角基部分开不远,伸向后外方,直或末端微向内弯。半牛的头骨角后部分长,角小而后伸,牙齿次高冠。在和政地区发现的秀丽半牛,其种名来自拉丁文,是“细瘦”的意思,这个种名表示其头骨比较窄细。秀丽半牛的角长而直,向外后方平伸,角基断面为前外边宽后内边窄的梯形,前上棱最锐利,前下棱次之,向上逐渐变为横向长的等边三角形。现生的水牛也具有三角形断面的角。