有些生物的形态看似在很长的演化历史中没有变化,仿佛演化出现了停滞。最近研究表明,这种现象不仅可能来自于维持性状不变的稳定选择,也可能来自于驱动性状变化、但方向频繁变化的定向选择。
从微生物到巨兽,地球生物千姿百态。在这迷人的多样性之中,有一个问题引发了演化生物学界的长期关注。那就是,如果演化导致了生物发生形态变化而具有多样性,为什么有些生物即使历经几亿年演化,似乎都不曾发生一点形态改变?近年来,不少研究证明形态的演化可以迅速发生——甚至是在代际之间发生。然而某些物种的演化似乎“停滞”了, “原地不动”。例如今天的腔棘鱼,与它们几亿年前的化石祖先几乎没有什么区别。
经过多年争论,演化生物学家们多用维持生物性状不变的稳定选择来解释这个悖论。直到近期,发表在《美国国家科学院院刊》(PNAS)的一项研究提出了另一种解释。该研究指出,那些看似形态不变的动物,其实可能是在短期的定向选择中不断发生性状变化;不过,放到更长的时间尺度里观察,定向选择频繁改变方向,以至于这些性状变化彼此抵消,导致了表面上的演化“停滞”。
悖论的源起
生物的形态与功能特征即所谓的“表型(phenotype)”。在生物演化的研究中,借助观察现存物种和化石记录中生物体多样的形态特征,我们能够描述表型在演化过程中的变化规律,并且试图解释这些表型变化和多样化的内在驱动机制。
具体来说,演化生物学家感兴趣的是表型演化的“节奏与模式(tempo and mode)”。表型演化的快慢规律,也就是“节奏”,给予了我们一窥演化驱动机制的宝贵信息,早在达尔文的研究和论述中就得到了关注。
按照达尔文的观点,生物演化主要是物种表型发生的可遗传的变异在自然选择作用驱动下逐代积累的过程。因此,在演化历史中,表型变化应当是缓慢而连续的“渐变” ;从共同祖先开始,不同物种逐渐分化的过程应该产生一些“中间类型”。然而,化石记录并没有证明这一推断。
对于物种表型演化的速率,达尔文在《物种起源》中写道:“不同纲以及不同属的物种,并没有依照同一速率或同一程度发生变化。在第三季最老的地层里,在很多灭绝了的类型中,还可以发现少数现生的贝类。……志留纪的海豆芽,与该属的现生种之间几无差异;然而,志留纪的绝大多数其他软体动物和所有的甲壳类,却已经发生了极大的变化。”(《物种起源》第十章“论生物在地史上的演替”)这些现象表明,表型演化在不同物种中存在速率差异。
这种差异显然需要进行机制上的解释。现生物种与化石记录形态相差无几,意味着演化树上的这一支系在长时间内缺乏表型的变化,即所谓的“演化静滞(stasis)”。例如,腔棘鱼类与四足动物的亲缘关系要比与鲤鱼等“真正的鱼”的关系更近,是肉鳍鱼类中最早分化出类群,绝大多数物种灭绝于六千多万年前;而现生的腔棘鱼类仅包含两个物种,其形态结构与化石记录中的白垩纪祖先几乎完全一样,是名副其实的“活化石”。那么在由自然选择主导的表型演化过程中,什么机制会导致我们观察到演化静滞呢?

↑ 西印度洋矛尾鱼形态

↑ 一块完整的腔棘鱼化石
达尔文认为,自然选择驱动下缓慢连续的表型变化加上“地质记录的不完整性”,可以解释表型演化的速率差异。从20世纪30年代开始,统计学的发展和孟德尔遗传学的再发现催生了 “新达尔文主义(neo-Darwinism,也叫现代综合论Modern Synthesis)”。以费舍尔(R. A. Fisher)、怀特(Sewall Wright)、霍尔丹(J. B. S. Haldane)、杜布赞斯基(Theodosius Dobzhansky)为代表的演化生物学家们为达尔文的学说提供了定量化的支持:他们使用基于遗传规律的概率模型,描述了物种表型如何通过生物种群内的自然选择作用驱动而发生演化的过程。
在新达尔文主义理论框架中,表型可以进行数量化:每一个表型的取值(或者几个表型取值的组合)都对应着一个生物体生存并留下可育后代的能力数值,即适合度(fitness)。适合度高的表型取值对生物体是有益的。
表型根据相关环境条件而受到几种不同的选择作用:发生线性的定向选择(directional selection)时,数值向一个方向变化对生物体生存和繁育后代是有益的,比如变大是有益的;而非线性的选择作用,包括数值“不大不小”最为有益的稳定选择(stablizing),以及数值变大和变小都更有益的歧化选择(disruptive selection)[1]。在生物种群繁衍一代的过程中,不同的选择作用导致种群中该表型的分布发生变化。
可想而知,定向选择可以使表型分布向某个方向偏移,逐代积累就会使整个物种的表型发生变化;而稳定选择则会使得物种的表型“原地不动”。因此,新达尔文主义认为,长时间持续的稳定选择可以解释我们在化石记录中观察到的演化静滞现象。
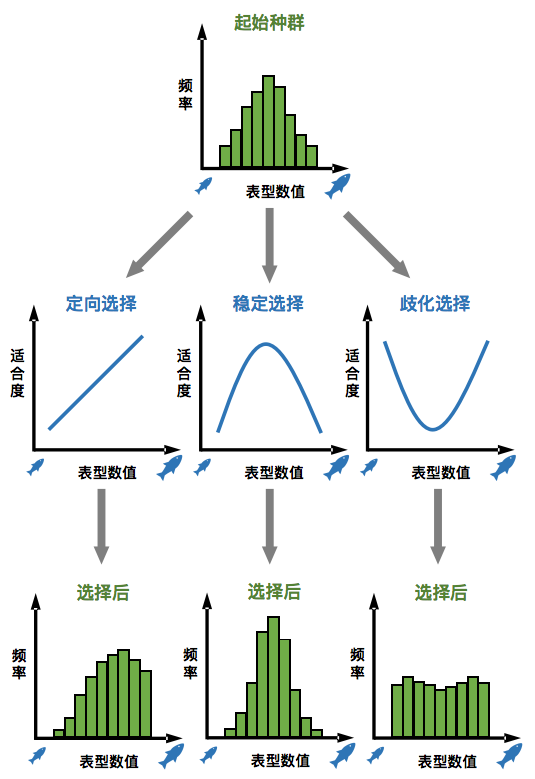
↑ 不同的选择模式及其对种群表型的影响(制图:卢平)
1972年,古生物学家史蒂芬·古尔德等人提出了“间断平衡论(Punctuated Equilibria)”来解释表型演化速率差异和演化静滞现象。间断平衡论认为,同一个物种在长时间的演化中,表型处于演化静滞,只是被物种形成事件所“打断”。古生物学家斯坦利(S. M. Stanley)将演化静滞归因于物种内不同种群的基因流动——只要没有形成互相存在生殖隔离的不同物种,这些种群就会因为彼此之间的基因流动,使得每个种群的表型维持在某个 “平均值” 附近,难以变化。而古尔德等提出发育限制来解释演化静滞:多细胞生物发育程序的精细复杂,也许限制了物种经历自然选择过程发生显著表型变化的可能性,这也有可能导致演化静滞。
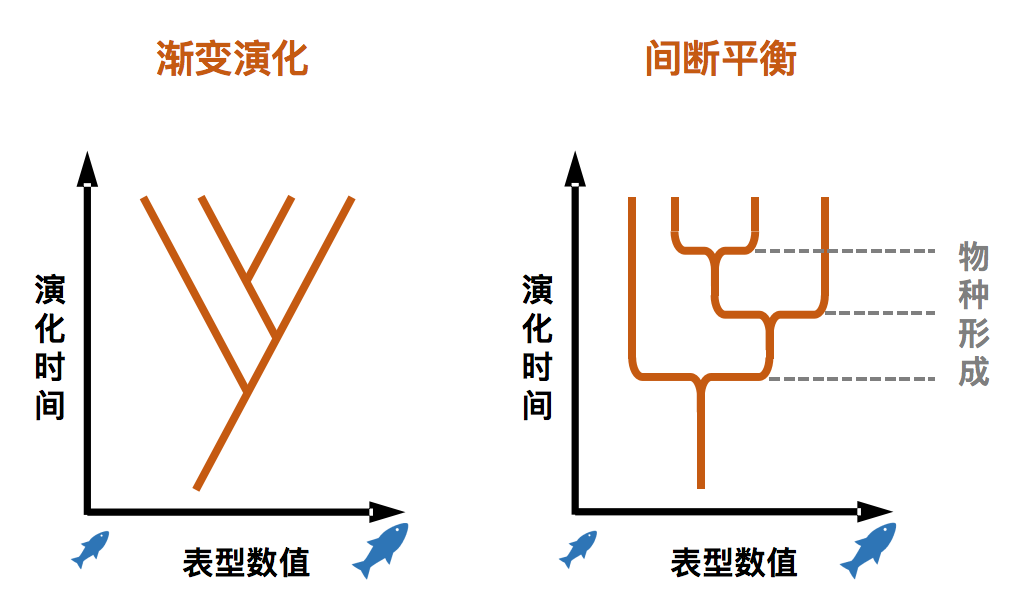
↑ 表型的渐变演化和间断平衡论(制图:卢平)
然而,新达尔文主义者们认为,间断平衡论无法对自然选择在表型演化和演化静滞中的作用构成挑战。查尔斯沃斯(Brian Charlesworth)和斯拉特金(Montgomery Slatkin)等人1982年的论述指出,发育限制难以完全解释演化静滞:家养动物在人工选择条件下表现出表型的巨大差异,例如不同家犬品系体型外表都相差很大;而野生物种中,也不乏桦尺蛾因为工业污染而在50年内颜色变化这种快速适应的案例。同时,表型差异也不总是跟物种形成事件相关,例如在相近的演化时间内,北美的小型雅罗鱼(minnow)类群演化出的物种数量是太阳鱼(sunfish)物种数量的数倍之多,然而两个类群表型变化程度相当,与物种形成事件的多少并无关联。因此 “渐变”式的表型演化并不罕见[2]。总之,新达尔文主义者相信,演化静滞的原因很可能主要是稳定选择。
但是稳定选择这一解释也面临困难。在现生物种中,令表型处于动态变化中的定向选择模式比比皆是,而维持表型保持在一个“最优值”的稳定选择模式很少被观察到,看起来不足以解释化石记录中常见的演化静滞现象。对此,有研究指出,当一个生物种群已经达到“最优值”、处于稳定选择时,稳定选择对种群中个体的适合度影响可能很小,因为大家都已经“差不多足够好”了,因此稳定选择可能在自然界中存在,但是难以从技术上测量验证[3]。演化静滞的普遍和稳定选择的少见,构成了表型演化领域所谓的“静滞悖论(Paradox of Stasis)”。
新近研究提出新机制
反思“静滞悖论”,这里的问题其实是 “节奏”与“模式”之间的关系。考虑一个适合度地形图(fitness landscape)模型:把地形图上X、Y两个轴想象成表型数量的两个维度,构成一个二维平面,也就是表型空间;平面上每个点的Z轴高度则表示相应表型的生物的适合度。生物种群对环境的适应可以看作是在这个地形图中向上攀登的过程。当我们考察这个地形图的一个局部,比如一个种群中所有表型的组合,发现这其中存在一个适合度的“山顶”,那么种群的表型应该受到稳定选择,趋向于集中在山顶;而当这个局部地形是一个斜坡,向着某个方向适合度不断增高,这就对应着定向选择,种群表型会偏离当前的分布,往那个方向变化。因此,当我们观察到演化静滞的“模式”,认为种群已经站上适合度的山顶,表型在每一代中都受到稳定选择保持不变;但是,实际的表型变化“节奏”,是否慢了下来呢?
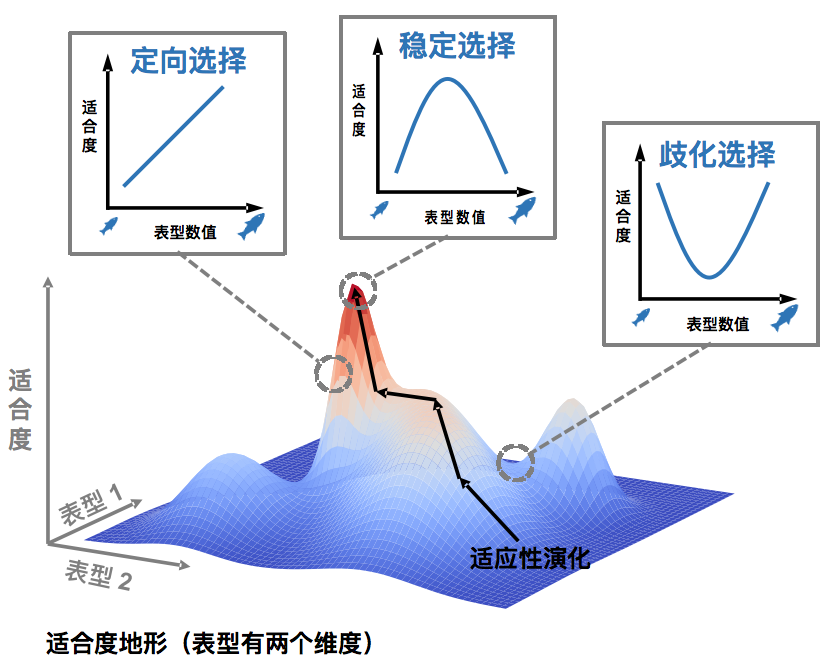
↑ 适合度地形图局部形状与选择作用的关系(制图:卢平)
不难想到,表型可能在某个固定值附近 “徘徊”,在每一代中都因为某些因素而向不同方向发生定向的变化;但是在较长的历史时期内,表型积累的净变化量仍然可以很小,看起来像是静滞。2016年,有针对大量化石记录的分析表明,在相同的时间长度内,静滞的表型和定向变化的表型实际上走过了相同长度的变化路程,只是前者的变化方向不定,因此净变化量远小于后者[4]。这也就从地质历史这一时间尺度上支持了“节奏”和“模式”并不一定匹配,静滞的表型仍在以相同的速率演化。
这样一来,表型的演化静滞就不必用稳定选择来解释了。那么表型在更小的时间尺度下的这种频繁变化,其驱动力又是什么呢?一个自然的假设是,种群每代(或者在延续多代的一个较短的时间段内)都受到朝向不同方向的定向选择,因而表型发生变化——确实,在自然界中,影响生物生存的环境因素肯定充满了变数,因而施加在表型上的自然选择作用也不可能一成不变。
2006年,华盛顿大学圣路易斯分校的乔纳森·罗索斯(Jonathan Losos)等人在《科学》杂志发表了一项研究[5],关注了分布于巴哈马群岛的一种安乐蜥Anolis sagrei。A. sagrei在没有天敌时,常常在地面上活动,当研究者们将另一种大型的地面捕食者Leiocephalus carinatus引入岛上时,A. sagrei面临新的生存挑战。一开始,后腿长的个体在地面上更容易逃脱捕食,但随着整个种群逐渐迁移到树上活动,后腿短的个体拥有更好的爬树能力。在为期一年的三次实地观测中,研究者们的确发现针对A. sagrei种群的自然选择从偏好腿长变成了偏好腿短。
短期内定向选择方向的变化,看起来是真实存在的。
2023年,罗索斯等研究者又发表了一项针对四个习性不同的安乐蜥物种的野外研究,直接探究了发生长时间演化静滞的表型在短时间内的动态[6]。研究者将持续两年半的研究分为五个时段,并在每个时段中测量不同个体的11种表型数据,例如前足长、头长等等。这五次测量结果表明,如果只看开头和结尾两个时间点,这些表型的适合度地形图存在一个 “山顶”,并且表型本身在这个最优值附近变化不大,像是在稳定选择下保持静滞。然而对每个单独的时间段,适合度地形图都是一溜“斜坡”,并且各个时间段的倾斜方向不同。例如在某些时间段,长腿有利于其生存,但在另外一些时间段,短腿更有竞争力。选择的方向和强度都有波动,而且并没有明显的变化规律。
这个结果意味着对于每一种安乐蜥来说,适合度地形图其实是随时间变化的,更像是“海景图(seascape)”而非地形图(landscape)——想象某个时刻海面波浪的形状,在下一时刻肯定会激荡变化。安乐蜥种群在每一代都受到某个方向一定程度的定向选择。一段时间内对生存最有利的表型,在另一段时间内可能并无生存优势甚至有害。由于方向不断变化,在更长的时间尺度上,适合度看起来有个累积的最优值,表型也会待在最优值附近,表现出演化静滞。
因此,这个研究表明,方向变动(fluctuating)的定向选择能够导致表型的演化静滞。

↑ 随时间变动的“海景图”导致方向变动的定向选择,也可能导致演化静滞(制图:卢平)
“静者动之基”,看似简单停滞的表型演化模式,却是复杂而动态的自然选择过程的结果。演化生物学研究由于关心的是历史,往往不得不用当前这个时间点观察到的现象去推断过去亿万年间发生的事件,并假设一些因素在时间尺度上保持恒定。自然科学研究的终极目标都是用简洁统一的规律和机制来解释研究对象的运行变化,然而没有足够细致精确的实验观测,我们可能难以发现和验证简洁规律底层的复杂机制。
事实上,在提到“表型演化在不同物种中的速率差异”时,达尔文就已经指出这“取决于很多复杂的偶发因素”,包括“变异的有利的性质”、“杂交的力度”、“繁殖的速率”、“缓慢变化的环境条件”等等。随着生物学大数据的不断积累和研究手段的不断进步,表型演化模式以及相关生物学因素——例如自然选择——在不同时间尺度上的复杂变化,仍会是研究者们追寻的有趣方向[7]。
参考文献:
[1] Futuyma, D.J. & Kirkpatrick, M. Evolution, xviii, 599 pages (Sinauer Associates, Inc., Publishers, Sunderland, Massachusetts, 2017).
[2] Charlesworth, B., Lande, R. & Slatkin, M. A Neo-Darwinian Commentary on Macroevolution. Evolution 36, 474-498 (1982).
[3] Haller, B.C. & Hendry, A.P. Solving the paradox of stasis: squashed stabilizing selection and the limits of detection. Evolution 68, 483-500 (2014).
[4] Voje, K.L. Tempo does not correlate with mode in the fossil record. Evolution 70, 2678-2689 (2016).
[5] Losos, J.B., Schoener, T.W., Langerhans, R.B. & Spiller, D.A. Rapid temporal reversal in predator-driven natural selection. Science 314, 1111 (2006).
[6] Stroud, J.T., Moore, M.P., Langerhans, R.B. & Losos, J.B. Fluctuating selection maintains distinct species phenotypes in an ecological community in the wild. Proc Natl Acad Sci U S A 120, e2222071120 (2023).
[7] Rolland, J. et al. Conceptual and empirical bridges between micro- and macroevolution. Nat Ecol Evol 7, 1181-1193 (2023).

来源:返朴公众号
作者:邹征廷(中国科学院动物研究所)
编辑:诸鹏飞
审核:盛捷