“大印度”问题的由来
东冈瓦纳大陆主要由印度、南极洲、澳大利亚和其他一些微大陆组成,在中侏罗世与非洲大陆分离。早白垩世(大约1亿年前),随着澳大利亚西北处新的洋中脊扩张,印度大陆从南半球中高纬度地区向北漂移,在晚白垩世时期到达热带-亚热带区域,最终在新生代早期(大约6000万年前)与欧亚大陆碰撞,这个过程及其后续的持续挤压导致了青藏高原的形成,也直接影响了全球的古地理、古洋流和古生态(Stampfli and Borel, 2002)。
在与欧亚大陆碰撞之前,印度大陆的大小要远远大于今天我们看到的印度大陆。科学家把印度克拉通北侧的主中央断裂(MCT)与雅鹿藏布缝合带之间的印度大陆向北延伸的部分称为“大印度”,代表着印度大陆北缘,目前南北向宽约200公里。“大印度”一部分俯冲至亚洲大陆之下,引起青藏高原的隆升,另一部分发生构造缩短。虽然已有古地磁、板块回路重建、地震、地形和构造变形定量恢复等地球物理学方法的研究,但“大印度”的大小依然存在较大争议,从300公里至3000公里不等(图1)。
“大印度”的大小对于理解多大、多厚的大陆岩石圈在什么样的牵引力下能发生深俯冲并形成今天我们看到的青藏高原、曾经多少印度大陆岩石圈发生了板内变形而缩短消失了、以及印度-亚洲大陆在什么时间什么地点发生了几次碰撞等板块构造重大科学问题至关重要。
板块回路重建的印度-欧亚大陆初始碰撞时期两个大陆之间的距离减去构造变形定量恢复的碰撞之后亚洲岩石圈的缩短量可以计算“大印度”北向延伸的范围。如果“大印度”是一个完整的大陆地壳。那么,越晚的印度-欧亚大陆初始碰撞所计算的“大印度”越小。大量地质学证据指示印度-欧亚大陆的初始碰撞发生于60 Ma左右。58 Ma时期(图2):板块回路重建的印度与欧亚大陆之间的距离(即汇聚距离)在西部和东部分别为3650 km和4550 km;基于构造变形定量恢复的新生代亚洲岩石圈缩短量在西部和东部分别为1050 km和1150 km(van Hinsbergen et al., 2019)。这两者的差值(即计算的“大印度”北向延伸范围)在西部和东部分别为2600 km和3400 km。构造变形定量恢复的特提斯喜马拉雅在碰撞之后的缩短量为600-1100 km。因此,采用60 Ma作为印度-欧亚大陆初始碰撞的时间,岩石圈缩短量与汇聚距离存在大约1500-2000 km南北向的赤字。
目前,科学家提出三种模型来试图解释岩石圈缩短量与汇聚距离之间如此巨大的赤字(图2)。模型一为陆陆碰撞模型,认为“大印度”上千公里的岩石圈在与欧亚大陆在60 Ma左右碰撞后被消耗而消失,未留下地质证据(图2A)。模型二与模型三提出60 Ma之后依然存在易俯冲的洋壳来解释印度与欧亚大陆之间大的汇聚距离。模型二为弧陆+陆陆两期碰撞模型,指出在印度与欧亚大陆之前存在蛇绿岩或者洋内岛弧,他们在60 Ma左右与印度大陆发生第一次碰撞,这时在欧亚大陆与印度大陆之间仍存在广阔的洋壳。印度与欧亚大陆的真正碰撞发生于40 Ma左右(图2B)。模型三为大印度洋伸展+双陆陆碰撞模型,指示在白垩纪晚期特提斯喜马拉雅微陆块从印度大陆分离,在“大印度”内部拉开一个洋盆(图2C),特提斯喜马拉雅微陆块与欧亚大陆碰撞之后,印度大陆才与欧亚大陆碰撞。
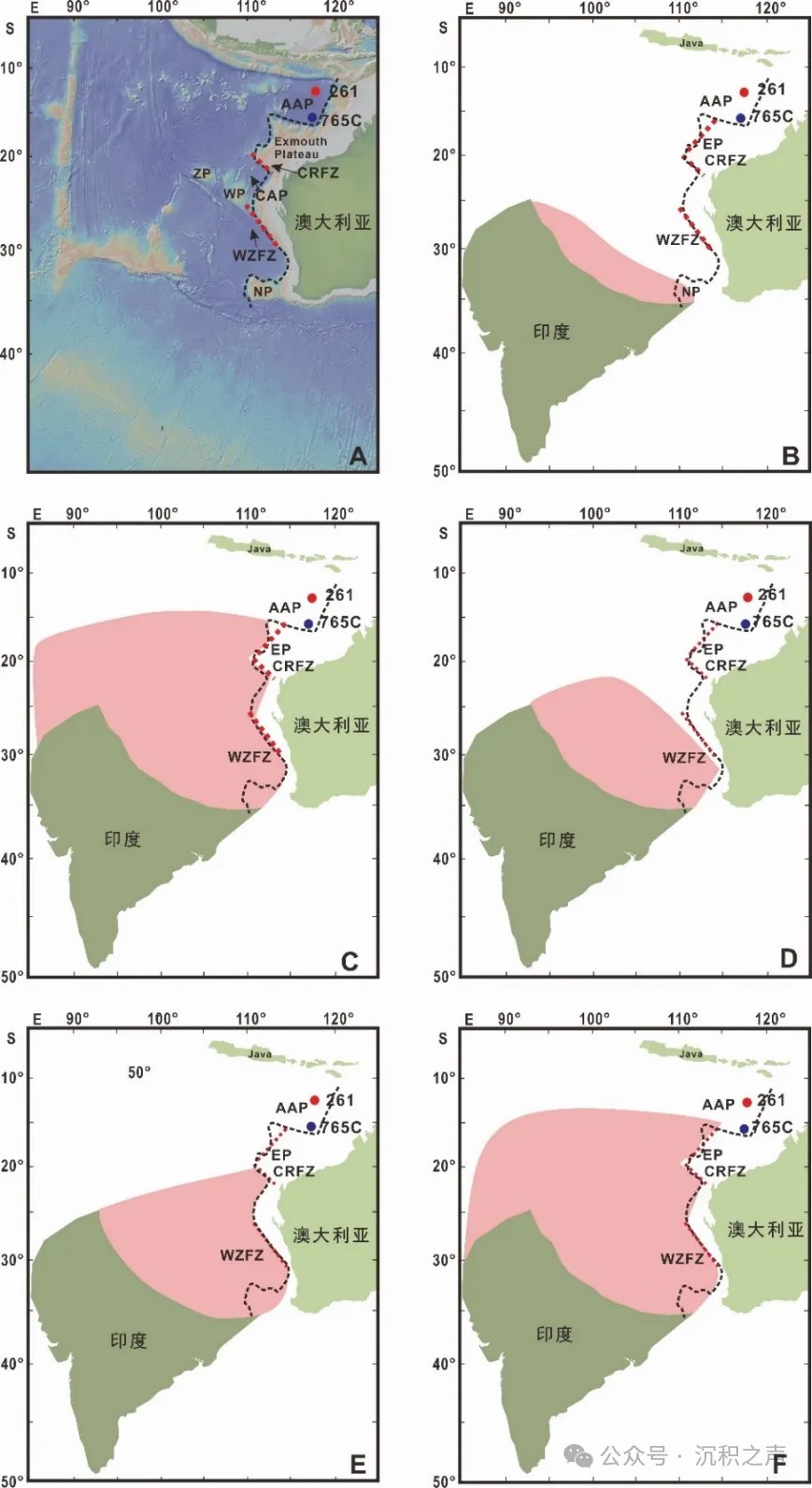
↑ 图1 重建“大印度”的北部边界与澳大利亚西北缘的相对位置(AAP-Argo深海平原,EP-Exmouth Plateau,CRFZ-Cape Range断裂带,WZFZ-Wallaby-Zenith断裂带):A.西北澳大利亚北缘地形图;B. Ratschbacher 等(1994);C. Lee and Lawver(1995);D. Ali and Aitchison(2005);E. Powell等(1988);F. Ingalls等(2016)。
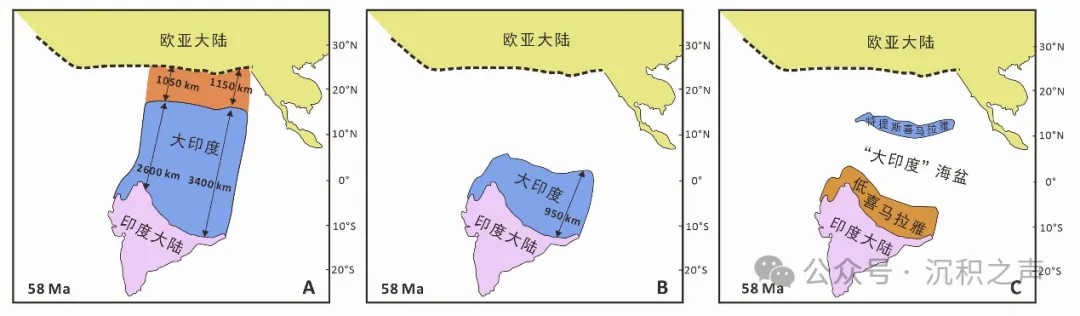
↑ 图2 不同“大印度”北向延伸与碰撞模型:A. 陆陆碰撞模型,58 Ma亚洲岩石圈的缩短量和较大的“大印度”;B. 弧陆+陆陆两期碰撞模型,较小的“大印度”与蛇绿岩或者洋内岛弧先碰撞;C. 大印度洋伸展+双陆陆碰撞模型,特提斯喜马拉雅微陆块先与欧亚大陆碰撞。
海洋微体生物–放射虫
在早白垩世早期,印度大陆与东冈瓦纳大陆开始裂解,此时印度大陆最北缘的边界位置是解决“大印度”北向延伸范围的关键(图3)。
放射虫是一种海洋原生动物,因含放射状的伪足而得名,单细胞,营海生漂浮生活。广布于世界所有海域,垂直分布可从表层至水深8000 m的海域。放射虫一般在热带真光层中多样性和丰富度最大,向两极和海洋深处递减,具有明显的垂直深度分带现象、纬向性、经向性和局域性分布特征(Boltovskoy, 2017)。放射虫个体较小(50–300 μm),种类繁多,演化迅速,是寒武纪以来生物地层学和生物古地理学研究中重要的门类之一。
放射虫包含等辐虫纲(Acantharea)、列足虫纲(Taxopodia)和多囊虫纲(Polycystinea)。多囊虫具有不易溶解的硅质骨骼,是放射虫中唯一能够于死亡后壳体在海底沉积物中得到很好保存的类群,又因其生态分布与海洋环境因子密切相关,从而在地层划分和对比有重要的作用,同时也可以应用于重建海表古温度等古海洋、古环境和古气候。因此,古生物学家或古海洋学家通常所说的放射虫(Radiolaria)一般指的就是多囊虫类放射虫(Polycystinea)。
放射虫的分布格局和群落结构受海洋物理性质(温度、盐度、深度、营养盐含量)和水文性质(上升流和洋流)等因素的影响控制。追溯和对比海相沉积中的放射虫动物群,成为恢复古洋流体系和大规模重建古海洋的重要方法。早白垩世早期,“大印度”的最北缘为广阔的特提斯洋南缘,在碳酸盐补偿深度(CCD)之下的深海区,由于其他钙质化石多被溶解,而硅质壳的放射虫可以被完好的保存下来,放射虫化石可以为该时期“大印度”北缘提供生物古地理方面的约束。
放射虫生物古地理
现代放射虫的生物地理分区为地质历史时期放射虫动物群应用于海洋古地理、古洋流和古环境的研究提供了重要的依据。根据不同的海洋生物建立的全球海洋分区模式为差别明显的带状的暖水区域(热带-亚热带)和冷水区域(两极)被两条带状的区域(过渡和亚两极)分开(Briggs, 1996)。这种分区模式与依据放射虫动物群建立的分区高度一致。早白垩世全球放射虫动物群存在明显的纬度分区现象。新特提斯洋之内的放射虫位于热带-亚热带地区(35°N– 40°S),呈现典型的中低纬度动物群面貌,被称之为特提斯动物群。南北半球中高纬度冷水区从晚侏罗世一直持续到晚白垩世,并含有不同于特提斯动物群的冷水动物群。Baumgartner et al. (2023)将早白垩世时期南半球的放射虫动物群分为新特提斯、泛大洋中部、东边界流、南方分区4个分区(图3)。新特提斯分区(Neotethyan Realm, NT)包含新特提斯洋内大型季风发育的区域和潘吉亚大陆内部的低纬度海盆(西特提斯洋、大西洋中部和原加勒比海)。放射虫动物群丰度和多样性高,常见有Baumgartner et al. (1995)中综述的465个种。泛大洋中部分区(Central Panthalassan Ream,CP)中的放射虫动物群与新特提斯分区中的高度相似。东边界流分区(Eastern Boundary Current Realm,EBC)位于潘吉亚大陆西部区域,包含较大的纬度范围(位于南北纬40°之间)。由于西风漂流到达大洋东部之后引起海流沿着大陆西缘向低纬度流动而造成温水和冷水放射虫动物群在该区域混合出现。例如:南极洲Longing Gap晚侏罗世至早白垩世早期的放射虫动物群混合了新特提斯洋内和南半球中高纬度种属(Kiessling et al., 1999)。南方分区(Austral Realm)的放射虫动物群以南半球中高纬度仅有的种属为主,主要包含Windaliinae 亚科中的 Praewindalia–Windalia 谱系、Fusitanellidae科和Parvicingulidae科的种属。
南半球早白垩世非特提斯冷水放射虫动物群的分布
在南极洲Weddell Sea (693B钻孔) (Ling and Lazarus, 1990)和Alexander Island(Holdsworth and Nell, 1992)、阿根廷南部的Patagonian Austral盆地(Hollis et al., 2009)、西太平洋595和596钻孔(Sutherland and Hollis, 2001)均发现有早白垩世的放射虫动物群,以非特提斯冷水种为主(图3)。这些动物群可能起源于南极洲Longing Gap (Kiessling et al., 1999) 和Alexander Island (Holdsworth and Nell, 1992) 以及新西兰的Waipapa地体(Aita and Spörli,1992)中晚侏罗世晚期的南半球非特提斯冷水动物群(Baumgartner et al., 2023)。
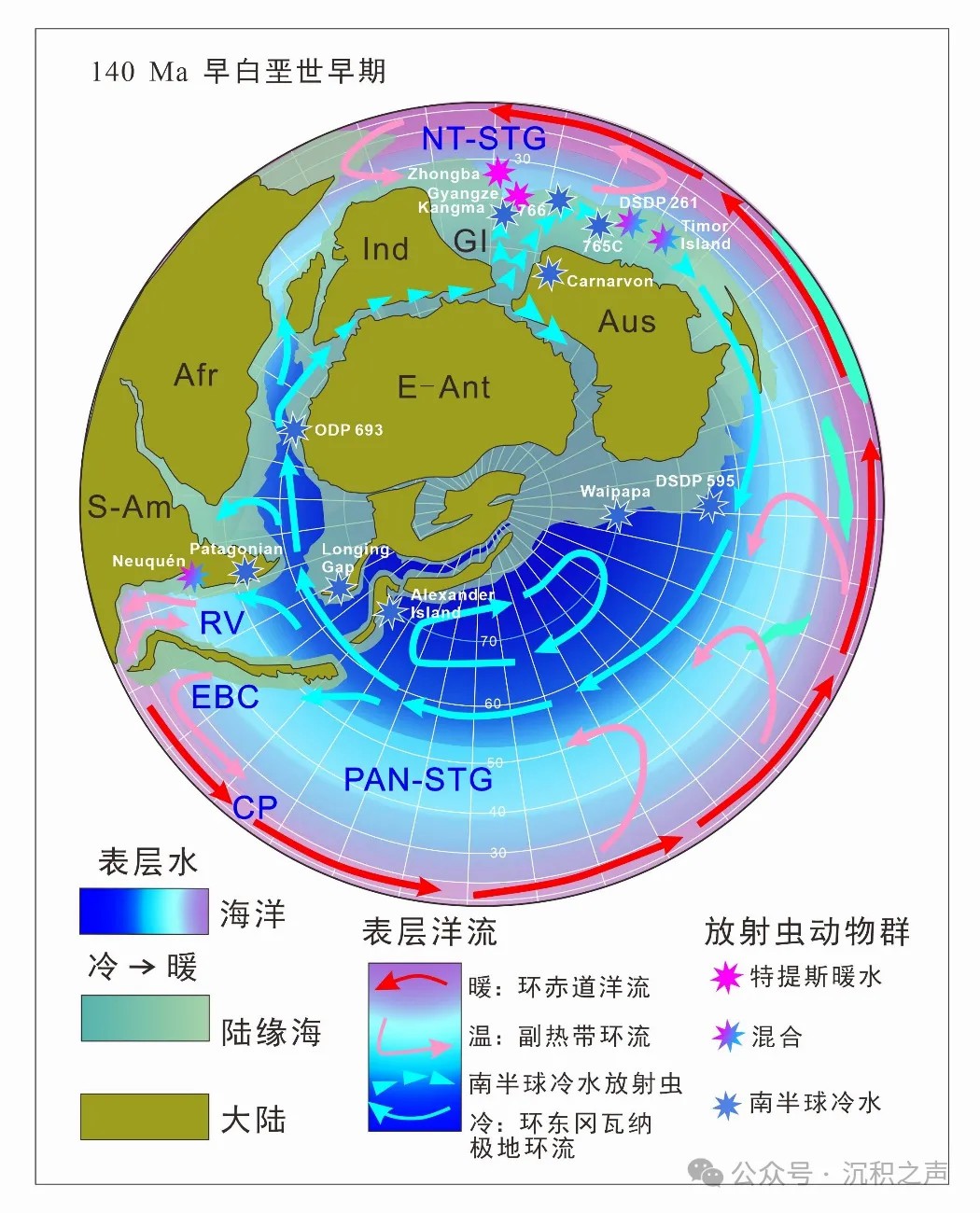
↑ 图3 早白垩世早期(140 Ma)东冈瓦纳古地理图(依据PANALESIS模型)。图中指示了古洋流、南半球放射虫古地理分区和已有的南半球冷水放射虫动物群报道及其扩散轨迹。
东冈瓦纳北缘早白垩世放射虫与“大印度”
在澳大利亚西北Argo深海盆地765站位的深海沉积中含有早白垩世的放射虫动物群(图4)。自晚贝里阿斯阶之后,黏土岩中的放射虫动物群多样性低,以“Crypto-Archaeo”型的罩笼虫等对生态耐受性强的世界种为主,缺乏典型的特提斯种属。在放射虫硅质岩中以南半球冷水种属为主,特提斯种属在欧特里夫阶/巴雷姆阶之后才开始逐渐出现,并逐渐增加。在西澳大利亚Carnarvon盆地(Ellis et al., 1993)中也发现有晚阿普特期至早阿尔必期南半球非特提斯冷水动物群。虽然已经有很多地区报道了南半球中高纬度特有的非特提斯冷水种,但是对于放射虫的分类学研究仍然存在不足。Baumgartner et al. (2023) 基于Argo深海盆地大洋钻探765站位中保存精美的放射虫化石,对早白垩世南半球放射虫动物群进行了系统分类和描述,共包含10个科(其中2个为新科:Fusitanellidae n. fam.和Windaliinae n. subfam.),18个属(其中7个为新属:Nodosphaera, Praewindalia, Pachycingula, Archaeotanella, Morchella, Fusitanella,和Argofusus)和55个种(其中30个为新种,14个未命名种)。
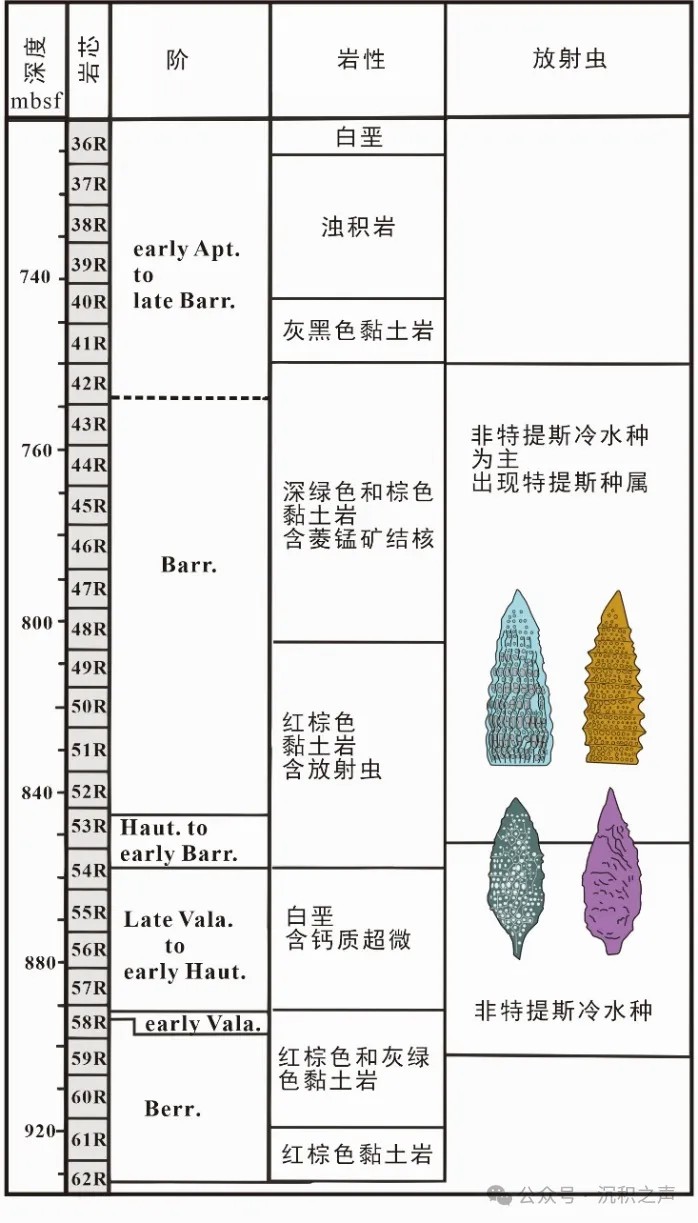
↑ 图4 Argo深海盆地大洋钻探765站位下白垩统深海沉积
伴随南极的冷水自南向北汇入,相近的古地理位置出现与结束南半球冷水放射虫动物群的时间(层位)应当一致,越北部的位置可能越晚出现南半球冷水放射虫动物群,更北部的位置可能不会受到南极冷水的影响而一直为特提斯暖水放射虫动物群。早白垩世放射虫动物群在东冈瓦纳北缘的分布特点可以为“大印度”的北部边界提供生物古地理方面的约束(图 5)。
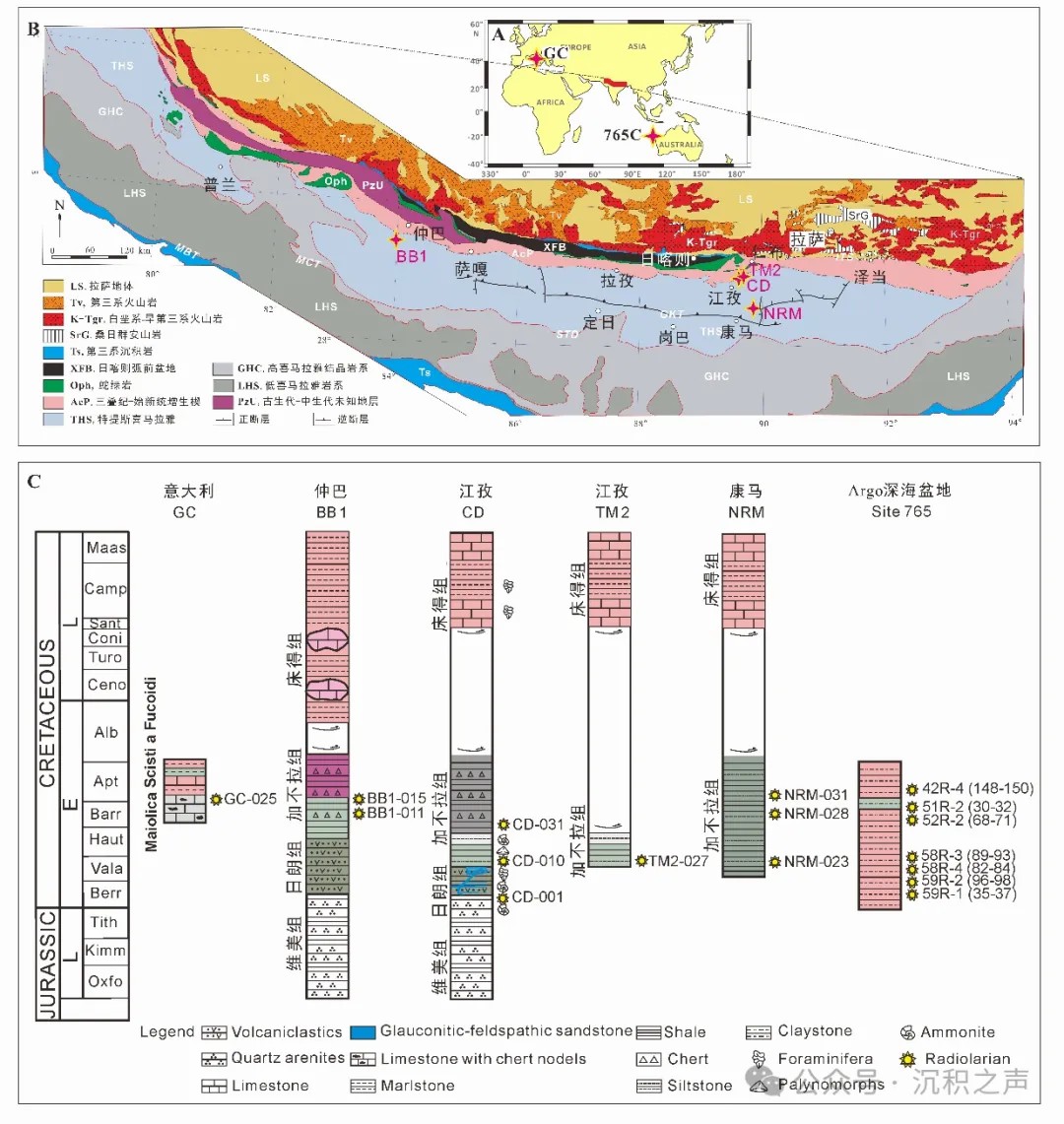
↑ 图5 研究剖面及采样层位A. 采样剖面分布图;B. 雅鲁藏布江缝合带地质简图;
C. 剖面柱状图及采样层位(GC=Gorgo a Cerbara剖面,BB1=巴巴扎东剖面,CD=床得剖面,TM2=托莫剖面,NRM=涅如麦剖面)
数量化理论方法最早在1950 年由林知已夫教授提出(Hayashi, 1950),它根据研究目的不同,在方法上可分为数量化理论I、II、III 和IV。“Hayashi数量化理论III”是一种可以同时对定性变量和定量变量进行处理的多元分析方法。该方法和因子分析、主成分分析等方法类似,都是用于分析样品或说明变量中起支配作用的主要因素或成分,并据以实现对样品或变量的分类。其基本原理是基于“0-1”属性判断矩阵的构建和向量值的计算,从而得到样本的得分。

↑ 图6 藏南江孜床得剖面和仲巴巴巴扎东剖面野外露头照片(A. 宏观照,B. CD-010,C. CD-031, D. BB1-011)。

↑ 图7 意大利Gorgo a Cerbara剖面野外露头照片(A. 宏观照,B. Marne a Fucoidi组多色泥灰岩,
C. Selli Level灰黑色页岩, D. Maiolica组灰白色灰岩夹灰褐色硅质岩条带,采样GC-025)。
为了定量化对比东冈瓦纳北缘早白垩世的放射虫动物群,Li et al. (2023)选取西藏南部3个点位(江孜、康马和仲巴)(图 6)所代表的印度大陆北缘海域、意大利(Gorgo a Cerbara剖面)(图7)所代表的西特提斯暖水域和Argo深海盆地(765站位)(图8)所代表的南半球中高纬度冷水域的6剖面共17个含放射虫的样品,进行统一的化学处理和鉴定,共包含贝里阿斯期-欧特里夫期的21个种和巴雷姆期-阿普特期的26个种。该方法将放射虫种属列表转换为“0-1”属性的二维判断矩阵,采用“Hayashi数量化理论III”的对矩阵进行运算得到放射虫不同种的得分,将放射虫动物群作为解释变量,将样品作为目标变量,实现对样品的分类(图 9)。

↑ 图8 Argo深海盆地含放射虫红色黏土岩采样层位
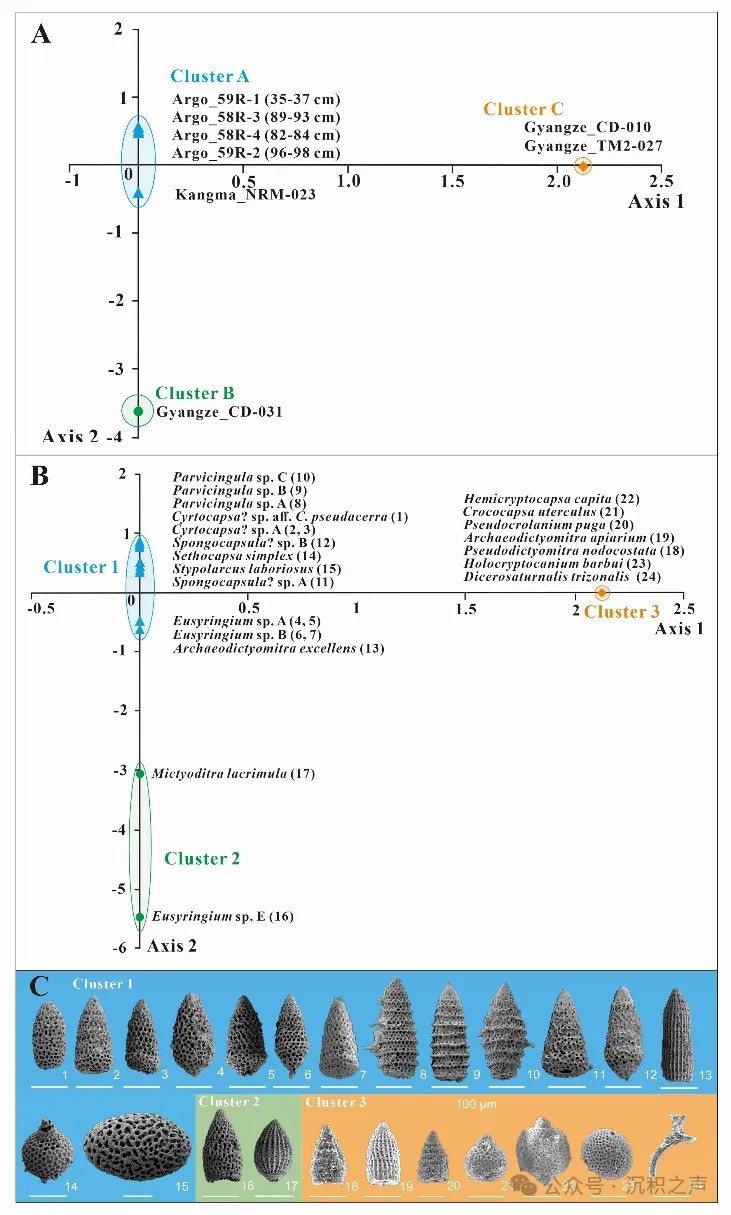
↑ 图9 基于“Hayashi数量化理论III”数值分析法分析的早白垩世放射虫动物群和样品散点图
研究表明(图10):(1)藏南部分地区出现的早白垩世南半球非特提斯冷水放射虫种属是由于印度大陆从东冈瓦纳大陆裂解,南极地区的冷水向北涌入;(2)同时期康马地区与Argo深海盆地765钻孔的放射虫均为南半球非特提斯冷水种属,指示相似的古地理位置;(3)江孜地区在130 Ma前后,放射虫动物群由特提斯暖水种为主转变为冷水种为主,指示在古地理上比Argo深海盆765钻孔位置更靠北;(4)仲巴地区在早白垩世时期一直以特提斯暖水动物群为主,指示了比江孜地区更靠北的古地理位置。
综合以上认识,早白垩世初期“大印度”最北缘可到达Exmouth 高原765钻孔更北部,南北宽大于2400 km。放射虫动物群的分布验证了印度大陆早白垩世时期的逆时针旋转,支持一个大的“大印度”模型。
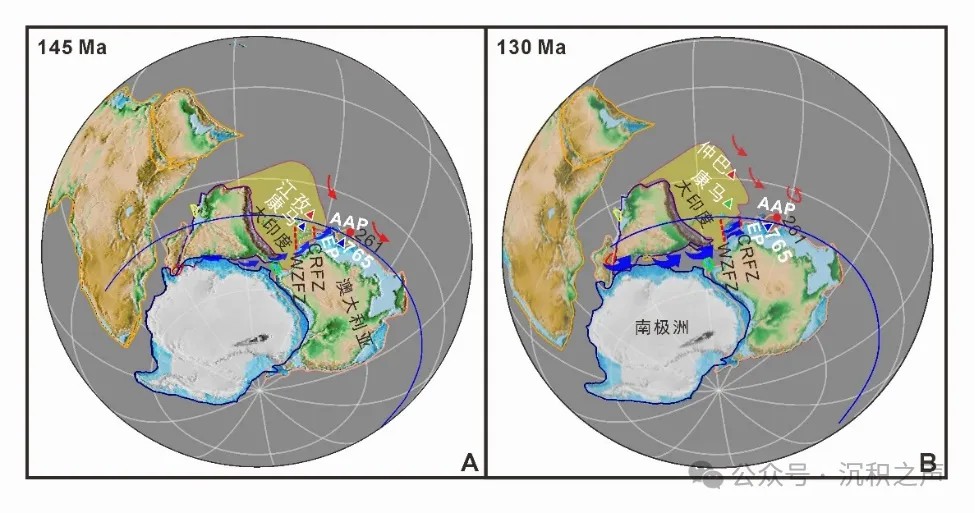
↑ 图10 基于放射虫动物群的早白垩世古地理重建图,蓝色箭头指示放射虫冷水动物群自南极向北扩散轨迹 (AAP-Argo 深海平原,CRFZ-Cape Range断裂带,WZFZ-Wallaby-Zenith 断裂带)。
来源:沉积之声
作者:李鑫
编辑:诸鹏飞
审核:盛捷