埃迪卡拉生物群(~5.74-5.39亿年前)记录了寒武纪大爆发之前复杂生物的演化过程。尽管许多埃迪卡拉型生物的系统发育位置尚未确定,但多种证据表明其中一些类型属于后生动物。厘清埃迪卡拉生物群的生物属性、系统发育位置及其不同组合的演替过程,能够让我们更好地理解后生动物的起源与早期演化。
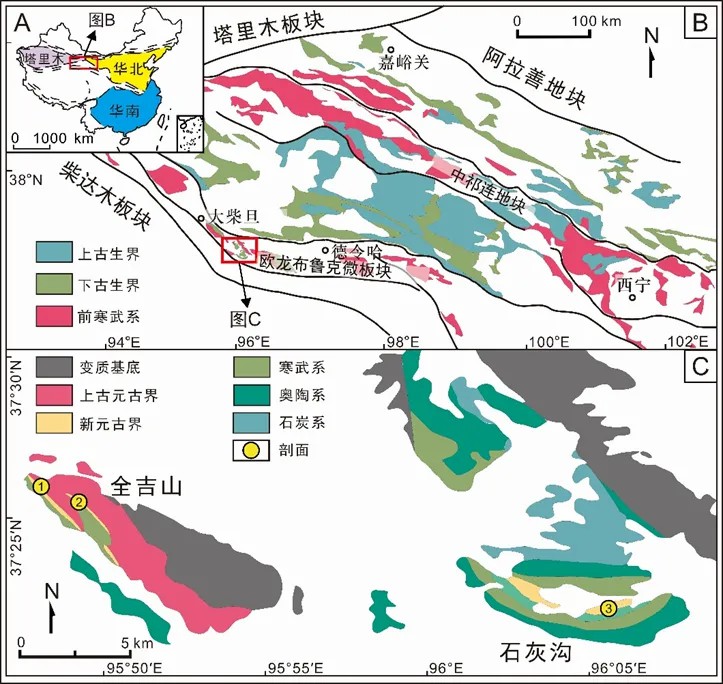
图1 柴达木盆地北缘欧龙布鲁克微板块区域地质图
为了弄清埃迪卡拉生物群内部的演化过程,研究者们基于全球化石分布的统计数据,通过定量的方法将埃迪卡拉生物群划分为三个主要组合,即阿瓦隆组合(~5.74-5.60亿年前)、白海组合(~5.60-5.50亿年前)和纳玛组合(~5.50-5.39亿年前)。然而,这些组合的形成在多大程度上受到环境、时间、古纬度或埋藏学因素的控制,仍然存在争议。因此,出现在多种岩性以及不同生物组合中的化石类型尤为重要,对它们进行研究有助于我们检验并进一步解释埃迪卡拉生物群的时空分布。

图2 中国科学院南京古生物所早期生命研究团队2024年在全吉山地区开展野外工作
近年来,在第二次青藏科考过程中,中国科学院南京地质古生物研究所早期生命研究团队在青藏高原北部柴达木盆地全吉山地区(图1;图2)皱节山组中发现了埃迪卡拉纪全吉山生物群,五年的野外工作采集了大量精美的埃迪卡拉型化石。该生物群包含两种极少在全球其他埃迪卡拉生物群产地共存的类型——恰尼虫(Charnia)和陕西迹(Shaanxilithes)。恰尼虫此前被认为是阿瓦隆组合中的代表分子,而陕西迹仅存在于埃迪卡拉纪末期(晚于5.50亿年),可能与纳玛组合是同时期的分子。因此,全吉山生物群可能代表了一个“过渡”型产地。这一发现为研究埃迪卡拉生物群的演化和时空分布提供了新的视角。

图3 全吉山生物群中的恰尼虫标本(比例尺均代表1 cm)
早期生命研究团队成员博士生吴承羲、助理研究员李光金、研究员庞科等人对来自全吉山生物群的恰尼虫化石进行了系统研究,为埃迪卡拉生物群中的叶状体这一重要类型提供了古生态以及埋藏学方面的新见解。相关研究成果近日发表在国际地学领域专业期刊《古地理,古气候,古生态》(Palaeogeography, Palaeoclimatology, Palaeoecology)上。

图4 全吉山生物群中的幼年期恰尼虫标本(A中比例尺代表5 cm;B–D中比例尺代表1 cm)
全吉山生物群所在的皱节山组岩性为细砂岩、粉砂岩和泥质粉砂岩互层。通过对来自该产地的69枚恰尼虫标本进行形态鉴别与测量,研究识别出了梅森恰尼虫(Charnia masoni)和细长恰尼虫(Charnia gracilis)(图3;图4),两个种主要通过一级分枝的形态以及分枝角度可以区分。此外,与世界范围内其他产地相比,来自全吉山生物群的恰尼虫标本整体分枝数量较少并且体型相对较小;部分标本的叶状体长度仅约1 cm(如图4B),一级分枝数量小于10,其大小以及一级分枝数量能够与前人报道的幼年期恰尼虫相对比。因此全吉山生物群的恰尼虫以小型个体为主,整体处于早期发育阶段,并且兼具部分处于更为幼年期的标本。

图5 来自全吉山A剖面皱节山组化石层位的沉积结构
(A–B)板状交错层理的野外露头照片。(C)平行层理的野外露头照片。(D–E)恰尼虫化石围岩的岩石薄片,含低角度板状交错层理(虚线框)。黑色箭头指示恰尼虫保存的层面。(E)中的白色箭头指示层理的截切(truncation)现象。黄色箭头指向岩层顶面。比例尺为10 mm。(F)为(E)中虚线框区域的放大图,显示了低角度板状交错层理的细节,以及磨圆较差的碎屑颗粒。比例尺为1 mm。(G)皱节山组下部产化石层位的整体岩性。(H–I)皱节山组下部可能的微生物诱导沉积构造(MISS构造),比例尺为5 cm。
对现代底栖群落的研究表明,叶状生物个体的小型化,可能有助于其生活在水动力较强的环境中。为了验证全吉山生物群的恰尼虫的小型化是否受环境因素影响,研究人员对化石所在层位的围岩进行薄片制作及观察,从而对恰尼虫的生活环境进行判断。根据野外剖面及薄片观察(图4),恰尼虫所在层位以泥质粉砂岩、细砂岩为主(图5A–C, G);结构成熟度高,成分成熟度低,碎屑颗粒磨圆度较差(图5F),表明其受水流作用影响小,整体处于低能环境。此外,在剖面露头上(图5A, B)以及岩石薄片中(图5D–F),均观察到了厘米级别的低角度板状交错纹层,其中既有直接位于化石层下方的交错纹层,也有位于化石层上方的层理截切现象。综合对岩性的观察以及沉积结构(平行层理、低角度板状交错纹层以及没有波痕构造)的识别,研究认为全吉山生物群的恰尼虫所在的古海洋环境处于下临滨带(lower shoreface)与过渡带(offshore transition)之间,水动力条件通常较低,可能偶尔有来自风暴事件的高能沉积物输入。

图6 全吉山生物群中三维保存的恰尼虫
(A)和(B)为同一恰尼虫标本的正反两面。在相邻的两个层面上,标本表现出相反的凹凸关系,说明该标本是三维保存的,叶状体上下两侧都在沉积物上形成了印痕。(C)恰尼虫标本以较高的凹凸程度保存,部分区域显著高于化石所在的层面。ne表示凹陷保存于岩层顶面,pe表示凸起保存于岩层顶面,ph表示凸起保存于岩层底面,nh表示凹陷保存于岩层底面。比例尺为1 cm。
埃迪卡拉生物群主要以二维印痕的形式保存,少数情况下也可以保存为三维模铸化石。通常情况下,叶状生物的固着器因为生活时扎根于沉积物中,更容易以三维的方式保存。全吉山生物群中包含了一些罕见的叶状体呈三维形式保存的标本。其中一个恰尼虫标本以三维模铸的形式保存(图6A–B),其所在微层从标本中部断裂,在相邻层面上呈现出凹凸关系相反的模铸,表明该微层的上下两面在成岩作用前均完成了模铸。这种三维模铸现象,也见于其他以凸起形式保存于岩层顶面或以凹陷形式保存于岩层底面的叶状体化石,表明它们很可能是三维模铸而非二维印痕保存(图7D)。完成叶状生物的三维模铸保存比二维印痕保存更具挑战性,因为标本需要在模铸完成前被沉积物完全掩埋,并保持外壁完整且不变形。在水动力条件较低的沉积环境中,三维模铸可能更容易实现。然而,沉积物是何时以及如何进入叶状体的内部空腔的,仍需进一步研究。此外,部分无固着器保存的恰尼虫标本可能在埋藏前经历过搬运,这也可能促成了其三维保存,相似的情况在纳玛生物群中近期有报道。
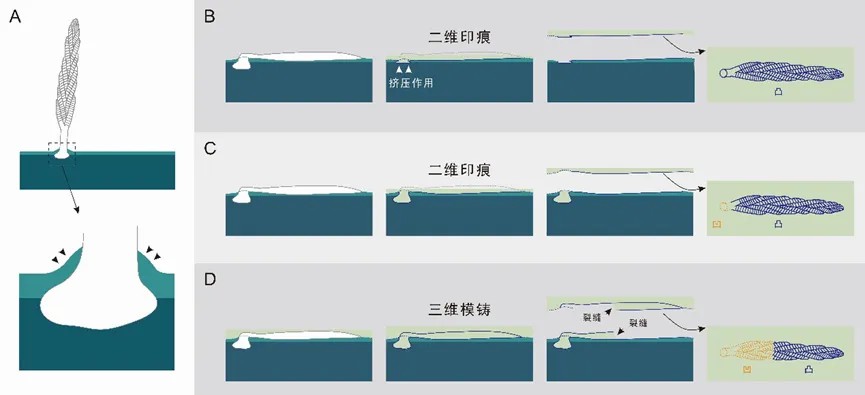
图7 具固着器的叶状生物二维和三维保存为化石的示意图
全吉山生物群中恰尼虫的发现,扩展了这一古老而又奇特的叶状生物的时空分布。恰尼虫与埃迪卡拉纪末期的潜在标准化石——陕西迹出现在同一层位,表明全吉山生物群中的恰尼虫可能是该化石类型最年轻的记录。综合世界范围内的化石记录,恰尼虫跨越了埃迪卡拉纪晚期约3000万年的地质历史,成功挺过了白海-纳玛组合之间的大灭绝事件,可出现在中临滨带(middle shoreface)到深海斜坡多种环境,表明这一叶状生物具有非常强的环境适应能力。
本研究得到了第二次青藏高原综合科学考察研究、国家自然科学基金委员会、中国科学院青年创新促进会、国家留学基金委、深时数字地球(Deep-time Digital Earth, DDE)国际大科学计划等项目的联合资助。
本次科普宣传与策划得到科技部国家重点研发计划青年科学家项目(2022YFF0802700)支持。
论文相关信息:
Wu, C., Liu, A.G., Liu, Y., Wang, X., Li, G.*, Qu, H., Huang, R., Qiu, M., Zheng, W., Sun, Y., Shi, H., Ouyang, Q., Wan, B., Chen, Z., Zhou, C., Yuan, X., Pang, K.* 2025. The Quanjishan Charnia assemblage from the northern Qaidam Basin, Tibetan Plateau, and implications for palaeoecology and taphonomy of Ediacaran fronds. Palaeogeography, Palaeoclimatology, Palaeoecology, 665: 112816.
参考文献:
[1] 吴承羲, 陈哲, 庞科, 王霄鹏, 万斌, 周传明, 袁训来. 2021. 三峡地区埃迪卡拉纪石板滩生物群定量古生物学和生态空间分析. 古生物学报, 60(1): 42–68.
[2] 吴承羲, 孙云鹏, 欧阳晴, 李光金, 周传明, 庞科. 2024. 柴达木盆地北缘元古宙地层的划分与对比评述. 地层学杂志, 48(1): 1–16.
[3] Pang K, Wu C, Sun Y, Ouyang Q, Yuan X, Shen B, Lang X, Wang R, Chen Z, Zhou C. 2021. New Ediacara-type fossils and late Ediacaran stratigraphy from the northern Qaidam Basin (China): Paleogeographic implications. Geology, 49(10): 1160–1164.
[4] Wu C, Pang K, Chen Z, Wang X, Zhou C, Wan B, Yuan X, Xiao S. 2024. The rangeomorph fossil Charnia from the Ediacaran Shibantan biota in the Yangtze Gorges area, South China. Journal of Paleontology, 98(2): 232–248.
[5] Chen Z, Zhou C, Xiao S, Wang W, Guan C, Hua H, Yuan X. 2014. New Ediacara fossils preserved in marine limestone and their ecological implications. Scientific reports, 4: 4108.
[6] Chen Z, Zhou C, Yuan X, Xiao S. 2019. Death march of a segmented and trilobate bilaterian elucidates early animal evolution. Nature, 573: 412–415.
[7] Fedonkin M A, Waggoner B M. 1997. The Late Precambrian fossil Kimberella is a mollusc-like bilaterian organism. Nature, 388(6645): 868–871.
[8] Waggoner B. 2003. The Ediacaran biotas in space and time. Integrative and Comparative Biology, 43(1): 104–113.
[9] Wang X, Liu A G, Chen Z, Wu C, Liu Y, Wan B, Pang K, Zhou C, Yuan X, Xiao S. 2024. A late-Ediacaran crown-group sponge animal. Nature, 630: 905–911.
[10] Xiao S, Chen Z, Pang K, Zhou C, Yuan X. 2020. The Shibantan Lagerstätte: insights into the Proterozoic–Phanerozoic transition. Journal of the Geological Society, 178(1): jgs2020–135.
编辑:诸鹏飞
审核:盛捷