原文来源:Adrián A D, Ben J W, Rochelle M S, et al., A geological timescale for bacterial evolution and oxygen adaptation.Science388,eadp1853(2025).
https://www.science.org/doi/10.1126/science.adp1853.

研究背景和科学问题
微生物在地球演化史上长期占据主导地位,但它们的化石记录极为稀少,这极大限制了对早期生命演化过程的认识,尤其是微生物类群起源时间的校准(即特定类群可能出现的最大年龄)。然而,细菌代谢活动通过同位素分馏或生物标志物在地球化学记录中留下的特征信号,为此提供了重要补充。地质历史上的大氧化事件(Great Oxidation Event, GOE,约24.3~23.3亿年前)是一个关键转折点,它由蓝细菌产氧光合作用的演化引发,并通过有机碳埋藏进一步促进大气氧含量上升。这一过程使生物圈从普遍缺氧状态转为有氧环境,进而推动了好氧代谢的适应性辐射。该研究利用大氧化事件事件后大气氧含量上升与好氧代谢扩散之间的时间关联,结合机器学习算法与系统发育调和分析(比较基因树与物种树的方法),重建了细菌祖先向好氧生活方式的转变历程,最终构建了细菌演化时间树并校准了关键分歧时间。
科学内涵
研究思路:
为了校准细菌演化树的时间框架,该研究创新性地利用大氧化事件作为关键时间节点,将其与好氧菌谱系的演化年龄相关联,从而引入了多个新的最大年龄校准点。研究采用贝叶斯分析方法,其核心假设是:好氧节点的演化时间通常不应早于大氧化事件,但在存在化石记录或序列分化证据支持的情况下,允许这些节点突破该时间限制。在方法学上,该研究将系统发育调和分析与机器学习技术相结合,以准确识别细菌演化树上厌氧到好氧生活方式的转变事件。通过整合全基因组数据,研究人员开发了一种能够从祖先基因组成的不完整信息中,稳健推断好氧与厌氧表型的方法。具体而言,该方法基于贝叶斯框架,要求只有在获得充分的化石或地球化学证据支持时,才能确定需氧节点的存在。通过将基因组分析与机器学习相结合,该研究实现了对细菌谱系从厌氧到需氧生活方式转变的精确推断。
核心发现:
这项研究基于包含1007种细菌的系统发育树分析,共鉴定出84次从厌氧到好氧的代谢转变事件。研究发现,这些转变主要发生在大氧化事件之后,其驱动力主要来自呼吸链相关基因和氧耐受基因的水平转移。
值得注意的是,虽然以大氧化事件作为时间校准点,但仍检测到至少3次转变早于该地质事件,这一发现表明,好氧呼吸作用可能在大气氧含量明显上升之前就已出现,并可能对蓝细菌产氧光合作用的进化产生了促进作用。
分子钟分析结果显示:细菌的最后共同祖先(the last bacterial common ancestor)可能出现在冥古宙至太古宙早期(44-39亿年前);细菌各门类的分化主要集中于太古宙至元古宙时期(25-18亿年前);而大多数细菌科的起源时间则可追溯至元古宙晚期(7.5-6亿年前),这一时间框架与陆生植物和动物门类的出现时期基本吻合(图1)。
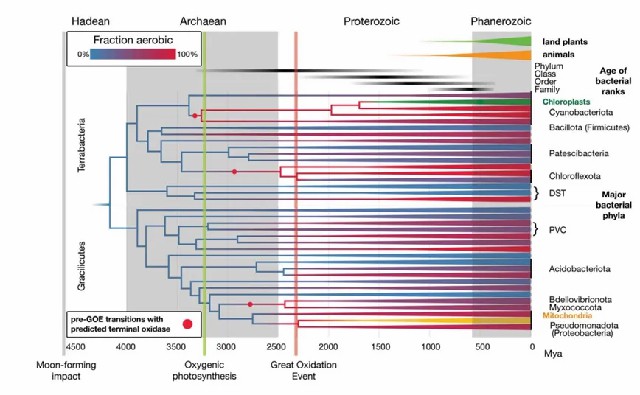
图1 细菌演化时间标定与氧适应历史重建的综合研究。该研究通过整合基因组数据、化石记录和地球化学证据,并将细菌的氧耐受性和好氧代谢与大氧化事件相关联,推断了细菌的演化时间树。图中蓝色表示厌氧状态,红色表示好氧状态,紫色深浅则表示现存细菌门类中好氧谱系的比例。研究还纳入了线粒体和质体数据,以利用更丰富的真核生物化石进行校准。此外,图中标注了陆生植物和动物的出现时间,以便进行时间对比。
综上,最早的好氧细菌在太古宙就已出现,比大氧化事件提前约9亿年,其中蓝细菌祖先的好氧代谢演化尤其关键,这一早期适应可能直接促进了产氧光合作用的发展(图2)。大多数细菌门类的祖先均为厌氧菌,其向好氧生活方式的转变主要发生在大氧化事件之后。分化时序显示,现存细菌门类的多样化始于太古宙至元古宙,而科级分化则普遍完成于显生宙之前(图3)。特别明显的是,大氧化事件后好氧菌谱系的辐射速率显著超越厌氧菌谱系,充分证明大气氧含量升高是驱动细菌演化的关键环境因子。
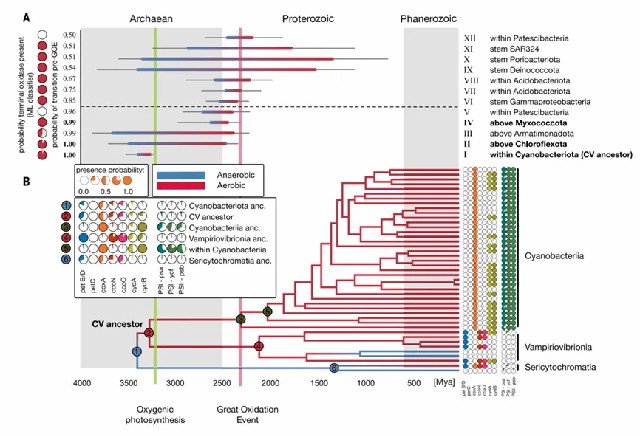
图2 有氧代谢早于大氧化事件(GOE)并可能促进产氧光合作用的演化。(A) 推断早于大氧化事件的有氧代谢转变时间范围(蓝色至红色渐变表示)。年龄区间从厌氧祖先谱系最老的5%分位点延伸至好氧后代谱系最年轻的5%分位点。12个转变事件的中位年龄早于大氧化事件(转变概率>0.5);其中5个转变事件在大氧化事件前发生的概率超过0.95,经最大似然分类器(ML)推断有3个携带HCO/cytbd末端氧化酶。(B) 蓝菌门(Cyanobacteriota)内有氧代谢转变与基因组成演化分析。研究推断有氧生活方式的转变发生在通向蓝菌纲(Cyanobacteriia)与Vampirovibrionia最后共同祖先的分支节点。产氧光合作用出现较晚,位于通向冠群蓝菌纲的基干谱系(节点3;即全群蓝菌纲内部)。虽然冠群蓝菌祖先极可能已具备产氧光合能力,但由于系统发育噪声影响下共历时分析的保守性,部分关键基因的存在概率仅为中等水平。XGBoost预测结果与现存及祖先蓝菌门中复合体III、IV基因的分布模式相符,为共同祖先(CV)已存在CoxA基因的假说提供了支持证据。各列数据显示光合作用相关基因的平均存在概率:PS I - Psa:光系统I初级反应中心;PS I - Ycf:光系统I组装相关;PS II - Psb:光系统II核心(产氧必需)。综合分析支持"演化出产氧光合作用的蓝菌谱系此前已具备有氧呼吸能力"的假说。绿色虚线(约3.23 Ga):存在光合作用化石与同位素证据的年代;红色虚线(约2.32 Ga):大氧化事件结束时间。背景交替色块表示不同地质时代。
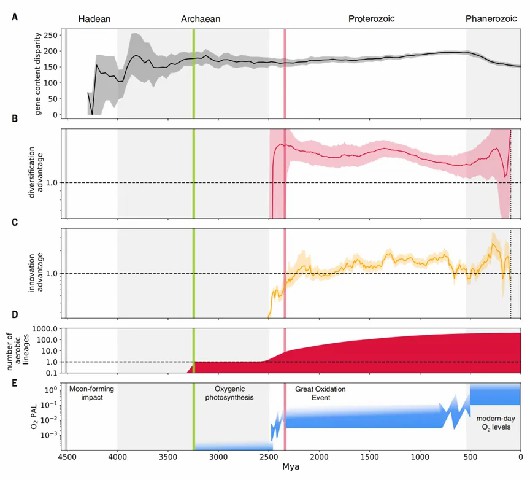
图3 地质时间尺度下的细菌演化历程。(A) 太古宙细菌遗传多样性的演化特征表现为基因内容差异度的快速上升。(B) 好氧与厌氧谱系的相对分化速率,结果表明自大氧化事件(GOE)以来,在大部分地质历史时期好氧菌都保持更高的分化速率。元古宙末期速率趋于平衡,随后好氧菌分化速率再次提升。虚线标示比值为1(即好氧与厌氧谱系分化速率相等)。(C) 好氧代谢通过水平基因转移(HGT)的扩散过程。自大氧化事件以来,厌氧向好氧的转变数量在大部分时期都超过逆向转变。曲线显示各时间段物种树上好氧/厌氧转变速率的比值。(D) 好氧谱系数量变化。(E) 大气氧含量变化。灰线(约45.2亿年前):MFI事件;绿线(约32.3亿年):产氧光合作用的化石与同位素证据;红线(约23.3亿年):大氧化事件结束。背景采用交替色块区分不同地质时代。
重要启示
环境与生命协同演化的新范式:该研究建立的细菌关键代谢特征演化时间框架(尤其针对厌氧与好氧代谢系统的起源和分化),使得微生物表型特征与地球化学历史的系统关联,地球化学证据与演化生物学理论的相互验证。这一创新框架为追溯其他重要性状在地质历史中的演化轨迹提供了新范式,开创了前寒武纪环境-生命协同演化研究的新方法体系。
编辑:诸鹏飞
审核:盛捷