

鳐类是鲨类的近亲 , 但外型上却和鲨类大不相同 , 牠们大多都是生活于海底 , 一共分作鳐目(Rajiformes), 锯鳐目(Pristiformes) , 鲼形目(Myliobatiformes)及电鳐目(Torpediniformes) , 只有锯鳐目(Pristiformes)及鳐目中的犁头鳐亚目(Rhinobatoidei)保留了鲨类的特征 。牠们是非常成功的鱼类 , 种类超过500种 , 世界各地海洋都可见鳐类的踪影 , 在这贴中会简略介绍一下鳐类的发展史 。
鳐类的先祖至今还是不明的 , 石炭纪的哥利鲨属Cooleyella牙齿表现了须鲨目及鳐类的特征 , 不过最早的鳐类化石记录是早侏罗纪 , 二者之间存有很大的空隙 。侏罗纪中期至晚期的原鳍棘鲨属Protospinax 和角鲨目 , 锯鲨目及鳐类关系非常密切 , 原鳍棘鲨属Protospinax的身体结构有着以上三者的特征 , 不过原鳍棘鲨属出现时间还是要较最早的鳐类来得晚 。鳐类 , 角鲨目及早侏罗纪的原鳍棘鲨属Protospinax关系密切 , 而鳐类较原鳍棘鲨属更早出现 , 这意味着鳐类可能于晚三迭纪已兴起 , 已知最古老的鳐类是早侏罗纪欧洲的Cyclarthrus , 只有离散的牙齿及刺剌化石被发现。
鳐目(Rajiformes)在基因上和鲨类很接近,尤其和猫鲨科更接近,牠一共分作尖犁头鳐科 (Rhynchobatidae) , 犁头鳐科(Rhinobatidae) , 鳐科(Rajidae) , 单鳍鳐科 (Arhynchobatidae) , 无鳍鳐科 (Anacanthobatidae)及拟鳐科 (Pseudorajidae)。单鳍鳐科(Arhynchobatidae),无鳍鳐科(Anacanthobatidae)及拟鳐科(Pseudorajidae)没有任何化石记录 , 不在这里详说。
早侏罗纪鳐类有着犁头鳐科(Rhinobatidae)的牙齿形态 , 牠们牙齿和其它新软骨鱼类(Neoselachians)不相同 , 大多不是被多层珐琅质覆盖 , 不过至少有两属是有着多层珐琅质覆盖 , Jurobatos 及Doliobatis牙齿就是被多层珐琅质覆盖 , 多层珐琅质结构看似是新软骨鱼类的原始特征 , 鳐类的牙齿珐琅质结构简单 , 牙齿结构和虎鲨属(Heterodontus)的臼齿化侧齿珐琅质相似 , 这看似鳐类牙齿有着先祖特征 , 一些史前鳐类也有着相似的牙齿结构 。
于法国 ,比利时及卢森堡发现了大量史前鳐类包括Toarcibatis multicristata, Toarcibatis brevicristata., Toarcibatis elongate , Toarcibatis alticarinata , Cristabatis crescentiformis , Cristabatis exundans及Doliobatis weisi 的牙齿化石,这些史前鳐类是属于早侏罗纪托阿尔阶(Toarcian) ,牠们牙齿有别于其它侏罗纪鳐类的,因此托阿尔鳐属Toarcibatis,Cristabatis及Doliobatis被划作古鳐科(Archaeobatidae)。
近来的数据显示尖犁头鳐属Rhynchobatids, 犁头鳐属Rhinobatids及鳐属Rajids是单源类群 , 团扇鳐属Platyrhinids则是鲼亚科Myliobatoidea的姐妹群 , 而犁头鳐属Rhinobatids的身体外型是属于原始特征 , 至于鲼亚科Myliobatoidea则是源于另一进化支。
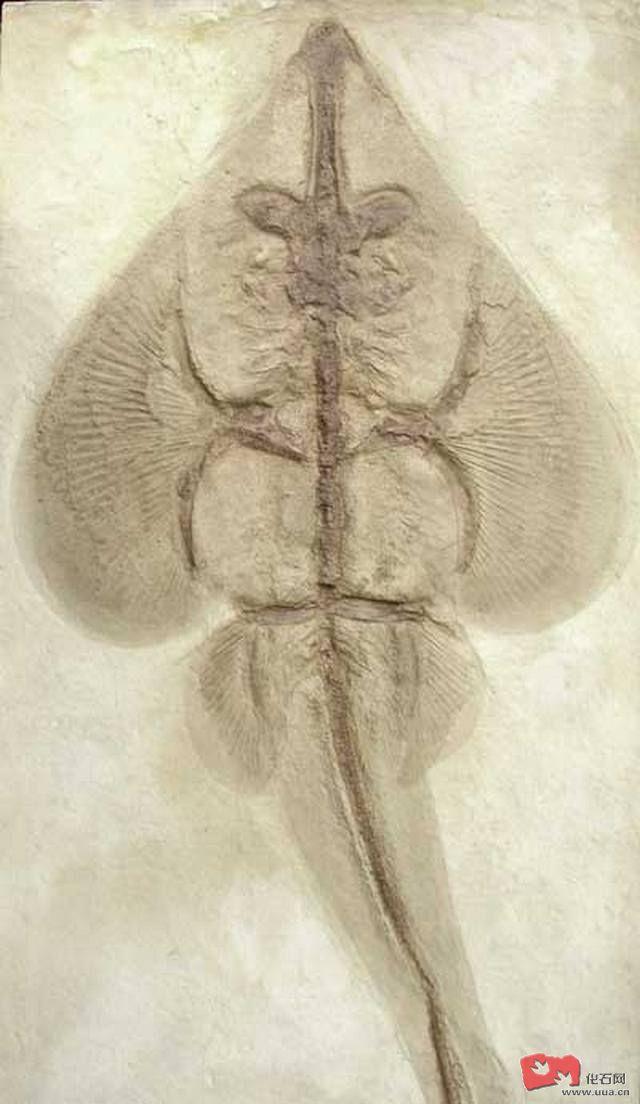
最古老有铰接的鳐类要追朔至晚侏罗纪(约一亿五千万年前) , 这些早期的鳐类化石于法国及德国等均有发现 , 比如箭鳐属Belemnobatis, 史帕斯鳐属Spathobatis及亚沙达鳐属Asterodermus , 牠们的形态已和今天的鳐类相似。亚沙达鳐属Asterodermus化石被发现于英国 , 法国及德国 。史帕斯鳐属Spathobatis及箭鳐属Belemnobatis这进化支要比锯鳐科(Pristidae)及尖犁头鳐科 (Rhynchobatidae)有更多先祖特征 , 比如鳍棘是被祖作原始特征 , 史帕斯鳐科(Spathobatidae)于晚早侏罗纪出现 , 属于鳐目(Rajiformes)最基层成员。一些史前鳐目成员如Hypsobatidae的系统位置还是不明的 , Hypsobatidae于坎帕阶(Campanian)至马斯特里赫特阶(Maastrichtian)存有化石记录。此外还有晚白垩纪的Germanobatos , Erguitaia , Ewingia , Tanoutia 及Turoniabatis 等一众史前鳐目成员归属也是存有疑问的。
Belemnobatis sismondae
犁头鳐科 (Rhinobatidae)于中生代有大量属系, 最古老的犁头鳐科(Rhinobatidae)成员是晚侏罗纪德国的Engaibatis schultzei。众多史前犁头鳐科属系中 , 值得一说的是角齿圆犁头鳐属Squatirhina , 牠的牙齿表现出鳐目及须鲨目的特征 。晚白垩纪至早第三纪的角齿圆犁头鳐属Squatirhina的归属存有疑问的 , Arambourg (1952) 及 Herman (1977)认为角齿圆犁头鳐属Squatirhina是和须鲨目关系要较密切 , 不过Cappetta于1987年认为角齿圆犁头鳐属Squatirhina应是划作犁头鳐科中。
现存的犁头鳐科有铲吻犁头鳐属(Aptychotrema) , 拟团扇鳐属(Platyrhinoidis) , 犁头鳐属(Rhinobatos) , 颗粒犁头鳐属(Scobatus) , 南犁头鳐属(Trygonorrhina)及强鳍鳐属(Zapteryx)六属 , 南犁头鳐属(Trygonorrhina)的化石记录只有北意大利始新世地层发现的Trygonorrhina dezignii , 犁头鳐属(Rhinobatos)的化石记录大多限于白垩纪时期 , 其它犁头鳐科属系的则没有任何化石记录。
犁头鳐属Rhinobatos是栖息于温暖的大陆架浅海海底 , 一共有26种 , 只有白点犁头鳐(Rhinobatos percellens)及白吻犁头鳐(Rhinobatos leucorhynchus)是生活于美国东岸(北卡罗莱纳州南部) , 环吻犁头鳐(Rhinobatos productus)及银点犁头鳐(Rhinobatos glaucostigma)则栖息于加利褔亚的水域 。
犁头鳐属Rhinobatos 于埃塞俄比亚晚侏罗纪托尔阶(Toarcian)地层已有化石记录 , 于白垩纪时期的北美 , 南美 , 非洲 , 亚洲及欧洲都有分布 , 于北美最早的犁头鳐属种化石记录是来自德克萨斯州阿尔布阶(Albian)地层 , Rhinobatos incertus 及Rhinobatos casieri是于白垩纪中期至晚期的北美很常见的种类。于黎巴嫩发现不少史前犁头鳐属化石包括Rhinobatos maronita, Rhinobatos hakelensis, Rhinobatos whitfieldi, Rhinobatos tenuirostris, Rhinobatos intermedius, Rhinobatos latus, Rhinobatos primarmatus ; 于当地也发现了形态酷似犁头鳐属的史前鳐类菱翼手鳐属种Rhombopterygia rajoides , 牠们都是属于晚白垩纪森诺曼阶及桑托阶 , 众多黎巴嫩发现的史前犁头鳐属种中以Rhinobatos maronita及Rhinobatos tenuirostris和现存的犁头鳐属种进化支形成有关连。于1991年古生物专家MAISEY认为白垩纪早期巴西的Rhinobatos beurleni和黎巴嫰的Rhinobatos hakelensis , Rhinobatos primarmatus及团扇鳐属Platyrhina有密切关系。Rhinobatos beurleni现被划作新属Iansan , Iansan beurleni是作为犁头鳐类 , 鳐目及鲼形目(Myliobatiformes)进化支中最基层的成员。
至于现存的属种中 , 只有环吻犁头鳐(Rhinobatos productus)于北美更新世地层有化石记录。
尖犁头鳐科仅有尖犁头鳐属一属,尖犁头鳐属(Rhynchobatus)栖息于东大西洋 , 印度洋及西太平洋的热带浅海 , 栖息深度达50米 , 牠们以甲壳类 , 软体动物 , 鱿类及小型鱼类为食。尖犁头鳐属(Rhynchobatus)一共有四种包括生活于南大西洋东部的非洲尖犁头鳐(Rhynchobatus luebberti) , 印度洋的及达尖犁头鳐(Rhynchobatus djiddensis) , 太平洋西部的澳大利亚尖犁头鳐(Rhynchobatus australiae) , 太平洋西北部的滑鼻尖犁头鳐(Rhynchobatus laevis) , 其中以及达尖犁头鳐(Rhynchobatus djiddensis)体型最大 , 全长能达3.1米。尖犁头鳐属(Rhynchobatus)最早于晚白垩纪出现 , 但晚白垩纪地层发现的尖犁头鳐属种化石还是没被定种 , 尖犁头鳐属(Rhynchobatus)于欧洲,非洲及日本都有化石记录 , 已被定种的史前属种包括始新世英国的奋森氏尖犁头鳐Rhynchobatus vincenti 及中新世北美的原尖犁头鳐 Rhynchobatus pristinus。
鳐科分布于各地浅海海岸及深海 , 于现在多达277种 , 牠们没有尾棘 , 身体表面长有许多微细棘状物及齿状物 , 雌雄个体存有性双态性 。最早期的鳐科成员包括森诺曼阶黎巴嫰的副鳐Rajorhina expansa (Pararaja expansa) , 晚森诺曼阶黎巴嫰的轮鳐属Cyclobatis及埃及晚森诺曼阶Bahariya组发现的Mafdetia tibniensis 。从副鳐Rajorhina expansa (Pararaja expansa)化石形态来看 , 副鳐Rajorhina expansa (Pararaja expansa)同时展现了鳐目及鲼形目(Myliobatiformes)的特征 ; Mafdetia tibniensis牙齿和现存的鳐科属种很不相同 。
副鳐Rajorhina expansa (Pararaja expansa)
轮鳐属Cyclobatis生活于晚白垩纪森诺曼阶的黎巴嫰 , 一共有四种包括Cyclobatis longicanudatus , Cyclobatis major , Cyclobatis oligodactylus , Cyclobatis tuberculatus 。轮鳐属Cyclobatis的外型使古生物专家最初把其划作电鳐科 , 后来于1980年则划归鳐科 。 轮鳐属Cyclobatis的牙齿形态和现存的鳐科成员大不相似 , 轮鳐属Cyclobatis有鞭状的短小尾部 , 尾部被二至三列短而狭幼棘状物覆盖 , 没有背鳍, 短小而非常狭窄的吻突都显示了明显的祖先特征 , 这些特征都使轮鳐属Cyclobatis和现存的鳐科成员难以作出比较 。尽管轮鳐属Cyclobatis骨盆带和现存的无鳍鳐属(Anacanthobatis)及肢鳐属(Cruriraja)的相似 , 不过牙齿形态的不同使古生物学家难以判断轮鳐属Cyclobatis及现存的无鳍鳐属(Anacanthobatis)及肢鳐属(Cruriraja)的关连。
瓦尔特鳐Walteraja exigua是生活于晚白垩纪马斯特里赫特阶(Maastrichtian) 瑞典的鳐科成员 , 牠的牙齿形态和现存的鳐科有显着相似 , 瓦尔特鳐Walteraja exigua是最早具备类似现存鳐科牙齿的早期鳐科成员 , 牠是生活于和暖浅海的鳐类。于1990年McEachran 及 Miyake假定鳐科在太平洋进化 , 不过瓦尔特鳐Walteraja exigua的发现支持了Long's于1994年发表的意见 ---鳐科的幅射进化中心位处北大西洋地带 。
其它已灭绝的鳐科成员还有早始新世比利时的Smithraja forestensis , Smithraja forestensis牙齿同时展现出鳐科及犁头鳐科的特征 , Smithraja是鳐目中最后灭绝的史前属系 , Smithraja forestensis和现存的鳐目成员没有任何祖属关系。
鳐属于晚白垩纪出现 , 晚桑托阶黎巴嫰的Raja davisi 仅只有细小牙齿及骨骼化石发现 , 该化石个体被认为是属于幼体 , Raja davisi的分类是存有疑问的 , 有专家认为Raja davisi和犁头鳐属Rhinobatos有较密切的关系 ; Raja sudhakari则是另一种被发现于印度马斯特里赫特阶(Maastrichtian)地层的鳐属成员 , Raja sudhakari牙齿和背棘鳐(Raja clavata)的相似 ; 至于德克萨斯州马斯特里赫特阶(Maastrichtian)地层发现的Raja farishi , 牠的牙齿和现存的鳐科成员大不相似 , Raja farishi可能需要划作他属。
于第三纪时期鳐属于西欧及非洲非常繁盛 , 第三纪属种有晚古新世尼日尼亚的Raja louisi , 早始新世摩洛哥的Raja praealba , 始新世前期英国的Raja harrisae , 渐新世比利时的Raja casieri , Raja cecilae , 中新世法国及瑞士的Raja gentili , 法国及荷兰的Raja olisiponensis, 上新世比利时的Raja antique , Raja clavata 。始新世前期英国的Raja harrisae牙齿和现存的圆身鳐(Raja cyclophora/ Atlantoraja cyclophora)的相似。
于日本中部瑞浪中新世地层发现了三种鳐科成员牙齿 , 其中二种鳐科牙齿化石类似同鳍鳐(Sympterygia bonapartii)雄鱼 , 第三种牙齿则类似渐新世比利时的Raja cecilae。
中新世法国及荷兰的Raja olisiponensis最初被误划归电鳐属(Narcine) , Raja olisiponensis于法国及意大利北部早上新世地层也有化石记录 。鳐科于早上新世的比利时非常繁盛 , 于比利时也发现了早上新世的Dipturus , 白魟属(Leucoraja)及鳐属的牙齿化石 , 于南加利褔亚上新世至更新世地层也发现了鳐科胸鳍及牙齿化石 , 值得一说的是现存的双斑鳐(Raja binoculata)于北美的中新世地层及上新世地层已有化石记录 , 背棘鳐(Raja clavata)于比利时上新世地层有化石发现 , 白令海深海鳐(Bathyraja interrupta/ Raja kincaidi )于北美的上新世地层也有化石记录 。
于众多史前鳐类中不得不谈硬尖犁头鳐科 Sclerorhynchidae ,硬尖犁头鳐科 Sclerorhynchidae 大约源于早白垩纪的巴列姆阶(Barremian) , 牙齿外型显示牠们和犁头鳐科 (Rhinobatidae)或锯鳐科(Pristidae)关系较近 , 硬尖犁头鳐科 Sclerorhynchidae现已被划归锯鳐目。
硬尖犁头鳐科 Sclerorhynchidae 一共多达24属,包括Ankistrorhynchus , Dalpiazia, Ischyrhiza, Kiestus, 硬尖犁头鳐属Sclerorhynchus ,裂根鳐属Schizorhiza , 帆锯鳐属Onchopristis , Onchosaurus , Pucapristis , Plicatopristis , Renpetia , Archingeayia , Baharipristis , Biropristis , 博罗地诺锯鳐属Borodinopristis , Celtipristis , Marckgrafia , 栉锯鳐属Ctenopristis , Ganopristis , 黎巴嫩锯鳐属Libanopristis , 微锯鳐属Micropristis , Ptychotrygon, Ptychotrygonoides ,大西洋锯鳐属Atlanticopristis及Texatrygon。硬尖犁头鳐科Sclerorhynchidae大多是生活于浅海及海湾底部的鳐类 , 属种全长大多不足1米。硬尖犁头鳐科 Sclerorhynchidae 中的一些类属如Celtipristis , Kiestus 及Ptychotrygon没有如锯鳐目般的剑吻锯齿化石发现 , 牠们仅只有口中的牙齿化石发现 , 尽管是被划作硬尖犁头鳐科 Sclerorhynchidae , 但是和真正的硬尖犁头鳐科 Sclerorhynchidae 成员关系还是存有争论 , 这些史前鳐类可能是属于别的进化枝。
于晚白垩纪北美东部海湾及内海就存活了硬尖犁头鳐科数个属系包括Ankistrorhynchus(桑托阶至坎帕阶) , 博罗地诺锯鳐属Borodinopristis(晚桑托阶至晚坎帕阶) , Ischyrhiza (土伦阶至马斯特里赫特阶), Kiestus(土仑阶至科尼亚克阶) , 帆锯鳐属Onchopristis(巴列姆阶至坎帕阶) , Onchosaurus(土仑阶至马斯特里赫特阶), Ptychotrygon(森诺曼阶至马斯特里赫特阶) , Pucapristis(晚桑托阶至马斯特里赫特阶) , 硬尖犁头鳐属Sclerorhynchus (森诺曼阶至坎帕阶) , 裂根鳐属Schizorhiza(坎帕阶晚期至马斯特里赫特阶)及Texatrygon(土仑阶至科尼亚克阶)。
博罗地诺锯鳐属Borodinopristis生存于晚桑托阶至晚坎帕阶北美 一共有二种包括Borodinopristis ackermani 及Borodinopristis schwimmeri , Borodinopristis ackermani是生存于热带浅海的小型鳐类 , 剑吻锯齿高度只有3mm。
Borodinopristis schwimmeri
Ischyrhiza于晚白垩纪的欧洲 , 非洲及美洲都很常见 , 一些属种吻突锯齿高达6 cm , 卵齿宽度达 7 mm , 于日本桑托阶至科尼亚克阶地层发现了新种Ischyrhiza iwakiensis 的锯齿相当粗大 , 长度达45 mm , 锯齿齿冠基部宽度达11.1 mm , 锯齿型态和北美的Ischyrhiza mira相似。
硬尖犁头鳐属Sclerorhynchus分布于晚白垩纪亚洲的黎巴嫰 , 欧洲古特斯海及北美都有化石发现 , 于黎巴嫰晚桑托阶地层发现了完整的Sclerorhynchus atavus骨骼化石 , Sclerorhynchus atavus全长1米。硬尖犁头鳐属Sclerorhynchus有长而呈鞭状的尾部 , 因此并不适合于远洋游弋 ,牠们肯定是底栖性鱼类。硬尖犁头鳐属Sclerorhynchus口腔中的卵齿非常细小 , 高度只有1.5 mm , 牙齿以甲壳类为食。
于巴西东北部发现了史前锯鳐类大西洋锯鳐Atlanticopristis equatorialis , 大西洋锯鳐Atlanticopristis equatorialis是早森诺曼阶(Cenomanian) 南大西洋浅海的硬尖犁头鳐科成员 , 牠可能会定期进入海口。从大西洋锯鳐Atlanticopristis equatorialis的尖吻锯齿形态来看 , 大西洋锯鳐Atlanticopristis equatorialis和帆锯鳐属Onchopristis的关系密切。
大西洋锯鳐Atlanticopristis equatorialis
被定种的帆锯鳐属Onchopristis一共有三种包括巴列姆阶至坎帕阶西欧及北美的Onchopristis dunklei , 阿普第阶至阿尔布阶德克萨斯州的古帆锯鳐Onchopristis praecursor及阿尔布阶至森诺曼阶北非的Onchopristis numidus , Onchopristis numidus吻突锯齿可达3吋长 , 可能是当时的大型锯鳐类 , 此外生活于马斯特里赫特阶北非Dalpiazia stromeri 锯齿也能长达53mm。于北美德克萨斯州发现的Onchosaurus pharao锯齿化石样本是发现众多史前锯鳐类中最大的 , Onchosaurus pharao锯齿化石于埃及 , 尼日尼亚 , 扎伊尔 , 巴西及安哥拉都有化石发现。
Onchosaurus sp
裂根鳐属Schizorhiza仅只有斯特莫裂根鳐Schizorhiza stromeri 一种 , 牠的化石于墨西哥及美国都有发现 , 生存于晚白垩纪坎帕阶(Campanian)晚期至马斯特里赫特阶(Maastrichtian), 全长1.5米。斯特莫裂根鳐Schizorhiza stromeri吻突的锯齿和其它锯鳐科成员及硬尖犁头鳐科 Sclerorhynchidae 的大不相同 , 吻突长有多达2百多颗排列紧密 , 细小锋利的针状锯齿 , 锯齿高达1~2 cm , 宽4~8 mm , 但卵齿(oral teeth)非常细小 , 高度只有1.5~2.5 mm 及宽 1 ~2 mm 。从斯特莫裂根鳐Schizorhiza stromeri的长吻来看, 牠们该不会以长吻挖掘泥沙的甲壳类及软体动物为食 , 反之会以长剑的小锯齿刺伤鱼类 , 此外斯特莫裂根鳐Schizorhiza stromeri有呈新月形的尾部 , 牠们会于陆缘海及浅海游弋猎食 , 而非底栖性鱼类。
Aramburg于1935年曾建议把锯鳐科分作锯鳐亚科Pristinae及Ganopristinae , Ganopristinae亚科的锯吻中的锯齿长有锯边 , 锯齿被珐琅质部分覆盖 , 马斯特里赫特阶北非 , 西欧 , 以色列及北美的Ganopristis锯齿能长达25 mm , 卵齿宽度能达35 mm 。Schaffer于1963年指出Pucapristis及帆锯鳐属Onchopristis和锯鳐属关系较密切 , Pucapristis的锯齿和锯鳐属的相似 , Pucapristis是生存于马斯特里赫特阶南美中部的锯鳐目成员。
现存的锯鳐目一共有两属 , 包括钝锯鳐属(Anoxypristis)及锯鳐属(Pristis) , 锯鳐类体型是仅次于前口蝠鲼属的鳐类 , 体型最细小的昆士兰锯鳐(Pristis clavata)雄鱼最大长3.06米 , 雌鱼最大长2.1米 ; 钝锯鳐(Anoxypristis cuspidata)最大可达3.5米 ; 最大的栉齿锯鳐(Pristis pectinata)平均长5.5米 , 最大达7.6米 ; 此外普通锯鳐(Pristis pristis)(全长可达5米) , 小齿锯鳐(Pristis microdon)(全长可达7米) , 大齿锯鳐(Pristis perotteti) (全长可达6.5米) 及后鳍锯鳐(Pristis zijsron) (全长可达7.3米)都是体型巨大的鳐类 , 其中大齿锯鳐(Pristis perotteti)重量达500至600公斤。
锯鳐类分布于非洲 , 澳洲及加勒比海等热带及亚热带海域 , 牠们也会于浅海湾及江河口出没 , 甚至河流也可见其踪影 , 例如生活于西非 , 加勒比海 , 美洲中部海岸及南美的大齿锯鳐(Pristis perotteti) , 牠们也会于尼加拉瓜湖(Lake Nicaragua)出现 ; 小齿锯鳐(Pristis microdon)及后鳍锯鳐(Pristis zijsron)分布于印度洋及西太平洋 , 昆士兰锯鳐(Pristis clavata)仅只分布于澳洲北部海岸 , 栉齿锯鳐(Pristis pectinata)于西大西洋的纽约至巴西 , 墨西哥湾及加勒比海 , 直布罗陀至非洲西南部海岸 , 印度洋及西太平洋 , 澳洲北部都可见其踪影 , 地中海可能也有其分布 , 普通锯鳐(Pristis pristis)则分布于东大西洋及地中海 ; 钝锯鳐(Anoxypristis cuspidata)从东非 , 也门 , 印度 , 泰国 , 印度尼西亚 , 澳洲 , 南中国海至黄海 , 日本西南部的海域都有分布。
锯鳐科有很悠久的历史 , 佩耶鳐属Peyeria就是目前发现的最古老的锯鳐科成员 , 牠是生活于晚白垩纪森诺曼阶北非的锯鳐类 , 佩耶鳐属Peyeria的长吻锯齿呈宽广的三角型 , 锯齿基部呈漏斗状 , 形态和始新世的锯鳐类很不相同 ; 其它锯鳐目成员都于始新世地层有化石发现 , 原锯鳐属Propristis是生存于始新世中期至中新世的欧洲 , 非洲及北美 , 被定种的仅只有始新世中期至渐新世的Propristis schweinfurthi , 牠的牙齿长达3 cm , 吻部锯齿没有珐琅质 , 基部有垂直沟及两个锯边 。原锯鳐属Propristis和现存的锯鳐属相似 , 都是生活于热带及亚热带的海岸 , 偶然会进入淡水。
钝锯鳐属(Anoxypristis)的化石记录大多来自始新世时期 , 属种有比利时及法国的Anoxypristis imhoffi(路德阶) , 西非南部的Anoxypristis ferinus , 尼日尼亚的Anoxypristis mucrodens , 法国的Anoxypristis priemi。于中新世的属种有刚果的Anoxypristis malembeensis及南卡罗莱纳州的Anoxypristis ensidens。
锯鳐属的化石记录要追溯至晚古新世时期, 最古老的属种是晚古新世塔内提阶(Thanetian)至始新世伊普雷斯阶摩洛哥的Pristis hamatus , 于始新世时期锯鳐属已取得广大的分布 , 属种有北美的Pristis agassizi , Pristis pickeringi , Pristis amblodon , 西欧,北美及北非的Pristis lathami , 非洲西部的Pristis olbrechtsi , 埃及的Pristis prosulcatus, 比利时的Pristis brevis , Pristis praecursor 及 Pristis propinquidens ; 于南极海岸也发现了锯鳐属未定种Pristis sp的节片化石。
Pristis lathami于始新世分布相当广泛 , 牠的吻突同时展现了钝锯鳐属(Anoxypristis)及锯鳐属的特征 , 于南卡罗莱纳州发现的Pristis lathami化石节片长达78 cm , 推测该吻突本来长119 cm , 全长推测达397 cm 至476 cm。于巴基斯坦发现的锯鳐属未定种(Pristis sp)吻突锯齿长达6cm , 于锯鳐属种中算相当大的 , 型态和普通锯鳐(Pristis pristis)及Pristis lathami的相似。于中新世的属种有早中新世至中新世中期法国的Pristis aquitanicus及中新世中期葡萄牙的大西洋锯鳐Pristis atlanticus , 现存的栉齿锯鳐(Pristis pectinata)于北美中新世及上新世地层都有化石记录。
南极始新世地层发现的锯鳐属未定种(Pristis sp)的节片化石
晚始新世至渐新世的锯鳐属新种Pristis fajumensis
鲼形目(Myliobatiformes)包括团扇鳐科(Platyrhinidae) , 梳板鳐科(Zanobatidae) , 六鳃魟科(Hexatrygonidae) , 近魟科(Plesiobatidae) , 扁魟科(Urolophidae) , 巨尾魟科(Urotrygonidae) , 魟科(Dasyatidae) , 江魟科(Potamotrygonidae) , 燕魟科(Gymnuridae) , 牛鼻鲼科(Rhinopteridae), 鲼科(Myliobatidae)及蝠鲼科(Mobulidae) , 大多成员脑部有较大的发展。
梳板鳐科是生活于东大西洋(非洲) , 于印度可能也有分布 , 该科只有梳板鳐属(Zanobatus) , 一共有梳板鳐(Zanobatus pusilla)及肖氏梳板鳐(Zanobatus schoenleinii)二种 , 外型和团扇鳐科(Platyrhinidae)相似 。团扇鳐科(Platyrhinidae)及梳板鳐科被认为是鲼科(Myliobatidae)的姐妹群 。
肖氏梳板鳐(Zanobatus schoenleinii)
团扇鳐科(Platyrhinidae)一共有二属三种 ,包括林氏团扇鳐(Platyrhina limboonkengi) , 中国团扇鳐(Platyrhina sinensis)及拟团扇鳐(Platyrhinoidis triseriata) , 团扇鳐科(Platyrhinidae)于亚洲 ,北美 , 墨西哥及加利褔亚邻近的北太平洋都有分布 , 于大陆架也可见其踪影。团扇鳐科(Platyrhinidae)有呈心形的头部 , 有像鲨尾的尾部, 尾前有二个背鳍。团扇鳐科(Platyrhinidae)的化石记录要追溯至白垩纪时期 , Pseudohypolophus , Myledaphus 及于意大利发现的特提斯鳐属Tethybatis都是已灭绝的团扇鳐科(Platyrhinidae)属系。
Pseudohypolophus 是生活于晚白垩纪北美内海的鳐类 , 牠的归属是存有疑问的 , 有专家认为Pseudohypolophus 应是划归犁头鳐科 ; 特提斯鳐Tethybatis selachoides是属于坎帕阶至马斯特里赫特阶 , 牠有粗而结实的鲨状尾部 , 较大的背鳍及尾鳍 , 特提斯鳐Tethybatis selachoides是团扇鳐科(Platyrhinidae)中的大型属种 。
Myledaphus bipartitus是生活于晚白垩纪 , 古新世至早始新世北美的河口及海湾, 是一种同时生活于河流及海洋的的鳐类 , 和现存的魟科(Dasyatidae)相似 , 能忍受高盐度。Myledaphus bipartitus全长不超过3呎 , 有着许多细小而平坦的牙齿以压碎甲壳类。Myledaphus属还有晚白垩纪乌兹别克斯坦的Myledaphus tritus。
团扇鳐属Platyrhina 则于晚白垩纪时期已存在 , 已灭绝的属种包括晚白垩纪意大利的Platyrhina bolcensis, Platyrhina egertoni 及早始新世美国的Platyrhina dockeryi。
魟科多达70种 , 大多是生活于热带及亚热带浅海 , 也有属种是生活于远洋 ,甚至有属种生活于湖泊河流 。 魟科成员会挖掘海底的蠕虫 , 甲壳类 , 软体动物 , 鱼类及头足类为食 。魟科有以下属种:
魟属(Dasyatis)一共有36种及5个未定种 , 分布于环球热带及温带海洋 , 从近岸水域至大陆坡上层 , 温带及热带河流湖泊都有分布 , 魟属于晚白垩纪时期已存在 ;
窄尾魟属(Himantura)一共有23种及4个未定种 , 分布于印度洋至太平洋的大陆架及海岛附近 , 西大西洋及东太平洋 , 于河流及湖泊都有分布 ;
长达4.2米的查菲窄尾魟(Himantura chaophraya)
萝卜魟属(Pastinachus)仅只有褶尾萝卜魟(Pastinachus sephen)一种 , 生活于热带印度洋至西太平洋大陆架 , 河流及湖泊 ;
Pteroplatytrygon仅只有Pteroplatytrygon violacea一种 , 一直被认为是属于魟属 , 分布于全球热带及温带的海洋。
条尾魟属(Taeniura)分布于热带及温带 , 东大西洋及印太平洋的大陆架及内陆架 ;
迈氏条尾魟(Taeniura meyeni)
沙粒魟属(Urogymnus)一共有二种 , 生活于东大西洋及印太平洋的大陆架 , 于西非的河流湖泊都有发现。
糙沙粒魟(Urogymnus asperrimus)
新魟属(Neotrygon)分布于印度洋至西太平洋, 最初还被划作魟属(Dasyatis)。余下的魟科成员还有双褶魟属(Amphotistius) 。
Neotrygon kuhlii
史前的魟科已有大量属系 , Coupatezia属牙齿化石于欧洲 , 北美及非洲都有发现 , Coupatezia属生活于晚白垩纪至始新世中期 , 牙齿类似现存的魟科 , 但牙根比例上则要大于现存的魟科属种 。和现存的魟科不同的是 , Coupatezia属牙齿齿冠唇面较宽。此外古生物学家Cappetta指出Coupatezia 属牙齿展现出性双态性 , 这点类似现存的魟科成员。
晚白垩纪马斯特里赫特阶的德克萨斯魟Texabatis corrugate有着类似魟科成员的牙齿 , 但牠的归属是存有疑问的。
德克萨斯魟Texabatis corrugate
于怀俄明州绿河的泥板岩上就可常看到完美的环棘魟Heliobatis radians化石 , 牠是属于始新世早期 , 环棘魟Heliobatis radians于很多方面类似现在的江魟科 (Potamotrygonidae) , 都是完全适应淡水的生活但不能于海洋生活 。当地也发现了另一种淡水鳐类Asterotrygon maloneyi的完整骨骼化石 , Asterotrygon maloneyi和扁魟属(Urolophus)有较密切的关系 , 牠的尾部 , 脑颅等都展现出许多鲼形目(Myliobatiformes)的近裔共征(synapomorphies)。于更早的时期就有异棘鲨类 , 弓鲨类中的滑齿鲨属(Lissodus)于淡水生活 , 直至现在只有约四十种软骨鱼类能适应淡水 ,但是排除淡水鳐类外 , 却只有非常少的种类能完全于淡水完成牠们的生命周期。
最古老的魟属成员是早白垩纪豪特里维阶(Hauterivian)英国的Dasyatis speetonensis , Dasyatis speetonensis的发现把鲼形目(Myliobatiformes)辐射进化时间推前至晚侏罗纪至早白垩纪 ; 魟属于晚白垩纪至第三纪世界各地都有许多化石记录 , 众多史前魟属成员中, 应该介绍一下两种魟属成员 : Dasyatis wochadunensis是生活于始新世伊普雷斯阶的英国 , 牠是一种体型巨大的魟属成员 , 于当地也发现了另一种魟科成员Hypolophodon sylvestris的牙齿化石 , Hypolophodon sylvestris是生活于古新世至早始新世的欧洲及西非 , 具有以磨碎食物为主的牙齿 , 牙齿高度能达7 mm。Hypolophodon sylvestris 牙齿有呈二裂的牙根及高而呈菱形的齿冠 , 牙齿形态和其它魟科成员大不相同。值得一说的是 ,当地也发现了猫鲨科新种Scyliorhinus biauriculatus , 锥齿鲨科的Synodontaspis striatus, Synodontaspis teretidens及扁鲨属种Squatina prima等生活于海湾的鲨类牙齿化石。
另一种要介绍的是非洲魟Dasyatis africana, 非洲魟Dasyatis africana最初于非洲古新世地层有化石记录 , 于古新世至上新世之间一直没有牠的化石记录 , 不过于埃塞俄比亚及肯尼亚的Turkana盆地上新世至更新世地层也发现了非洲魟Dasyatis africana的化石 , 非洲魟Dasyatis africana于190万年前从印度洋进入Turkana盆地形成淡水物种 , 这时候鄂嫩河(Onon River )流进主要湖泊及通过东南部 , 而图尔卡纳河把Turkana盆地及印度洋连接一起 。非洲魟Dasyatis africana于当地河流繁荣超过50万年 , 于130万年前才完全灭绝 , 非洲魟Dasyatis africana恐伯是魟属中最长寿的物种 , 从古新世一直坚持至更新世时期并演变成淡水物种 , 没有甚么物种能有非洲魟Dasyatis africana般的长寿 , 不能不说是一种奇迹!
魟属于中新世时期的大西洋东西两边都显得极具多样性 , Cappetta于1970年列出六种于中新世时期分布于大西洋东部的魟属成员包括Dasyatis cavernosa , Dasyatis probsti , Dasyatis serralheiroi , Dasyatis rugosa , Dasyatis minuta 及 Dasyatis delfortriei 。此外现存的美洲魟(Dasyatis americana)及粗尾魟(Dasyatis centroura)于北卡罗莱纳州早中新世地层都有化石记录 , 至于萨氏魟(Dasyatis sayi)则于北卡罗莱纳州上新世地层才有化石记录。此外 , 于加利褔亚鲨齿山中新世地层也发现类似萨宾河魟(Dasyatis sabina)的魟属牙齿化石。
江魟科(Potamotrygonidae)主要生活于南美淡水 , 有三属包括副江魟属(Paratrygon) , 近江魟属(Plesiotrygon)及江魟属(Potamotrygon) , 一共多达二十种。近江魟属(Plesiotrygon)及江魟属(Potamotrygon)都有呈尖角的软骨 , 只有副江魟属(Paratrygon)是没有。江魟科长有直肠腺 (rectal gland)以分泌盐份 , 也能减少及降低尿素浓度 , 这些都是适应淡水生活的证据。条尾魟属(Taeniura)有三个海生种类归属上是存有疑问的 , 这三种都是分布于大西洋东部 , 地中海及印度洋至西太平洋 ; 此外窄尾魟属(Himantura)中有两个海生属种的归属也是存有疑问的 , 包括生活于南美北部及美洲中部邻近太平洋一带, 长达1.5米的太平洋窄尾魟(Himantura pacifica) , 北美南部及南美北部邻近大西洋一带, 长达2米的施氏窄尾魟(Himantura schmardae) , 尽管牠们是放置于鳐科中 , 但还是需要更多的研究以了解牠们的关系。 Brito及Deynat于2007年假设江魟科于晚白垩纪或早第三纪时期已兴起 , 不过江魟科Potamotrygonidae于中新世时期才有化石记录。有专家认为江魟科Potamotrygonidae是源于太平洋 , 牠们的先祖可能是类似扁魟属(Urolophus)并且于古新世至中新世时期已局限于淡水 。
巨尾魟科(Urotrygonidae)分布于东太平洋及西大西洋的大陆架 , 至于近魟科(Plesiobatidae) , 扁魟科(Urolophidae)则分布于西太平洋的大陆架及岛坡 , 近魟科 Plesiobatidae只有一种---达氏近魟Plesiobatis daviesi , 最大可达2.7米。达氏近魟Plesiobatis daviesi分布于南非至夏威夷一带的印度洋至西太平洋 , 大陆架及岛坡都有其踪影。扁魟科(Urolophidae)包括鹞扁魟属(Trygonoptera) , 大尾扁魟属(Urobatis)及扁魟属(Urolophus) , 扁魟属(Urolophus)于始新世时期已留下化石记录 , 已灭绝的属种包括始新世早期意大利的Urolophus crassicouda 。带纹扁魟(Urolophus cruciatus)于北美晩始新世地层已有化石记录 , 哈氏大尾扁魟(Urobatis halleri)则于北美更新世地层才有化石记录。
鹞扁魟属种Trygonoptera testacea
哈氏大尾扁魟(Urobatis halleri)
海滨扁魟(Urolophus orarius)
带纹扁魟(Urolophus cruciatus)
达氏近魟Plesiobatis daviesi
六鳃魟科(Hexatrygonidae)是燕魟科(Gymnuridae)及鲼科(Myliobatidae)的姐妹组群 , 生活于南非至夏威夷一带的印度洋至西太平洋的大陆架及岛坡 , 该科只有六鳃魟属 。
比氏六鳃魟(Hexatrygon bickelli)
燕魟科(Gymnuridae)是分布于环球岛架及大陆架的热带及亚热带水域 , 牠一共有两属包括鸢魟属(Aetoplatea)及燕魟属(Gymnura) , 鸢魟属(Aetoplatea) 及大多燕魟属(Gymnura)分布于印度洋及西太平洋 , 燕魟属(Gymnura)和鸢魟属(Aetoplatea)不同之处在于燕魟属(Gymnura)没有背鳍 , 现存的燕魟属(Gymnura)一共有十二种 , 其中的大燕魟(Gymnura altavela)及小尾燕魟(Gymnura micrura)分布于于北太西洋西部 。燕魟科属种属底栖型鳐类 , 以鱼类 , 甲壳类及浮游生物为食 , 属种中小尾燕魟(Gymnura micrura)体宽只有1.3米 , 密斑燕魟(Gymnura crebripunctata)更细小 , 体宽只有31厘米 , 体型最大的大燕魟(Gymnura altavela)体宽则可达4米。
燕魟科(Gymnuridae)最早于晚白垩纪时期已出现 , 最古老的燕魟属种是晚白垩纪森诺曼阶埃及的Gymnura laterialata , 其它已灭绝的属种有古新世晚期摩洛哥的Gymnura delpiti , 比利时也发现了晚始新世的Gymnura grootaerti及渐新世鲁培勒阶(rupelian)的Gymnura hovestadti。燕魟科还有已灭绝的Subathunura casieri (早始新世印度) , Jacquhermania duponti 及Ouledia sigei , Jacquhermania duponti是被发现于维吉尼亚州及比利时始新世中期地层 , 牙齿高度可达2.5 mm , Ouledia sigei则是始新世摩洛哥的燕魟科成员 , 具体资料不详。
菱鳐科(Rhombodontidae)是晚白垩纪马斯特里赫特阶(Maastrichtian)最常见的鲼形目(Myliobatiformes)成员 , 一共有两属包括玻利维亚的Pucabatis(仅只有Pucabatis hoffstetleri 一种) 及菱鳐属Rhombodus , 菱鳐属Rhombodus的化石于欧洲 , 近东 , 非洲及美洲都有发现 , 菱鳐属Rhombodus的牙齿呈菱角型 , 有像牛鼻鲼属的齿板 , 但是菱鳐属牙根是二裂的 , 牛鼻鲼属的牙根是多裂的。Rhombodus levis牙齿于新泽西州及北卡罗莱纳州晚白垩纪地层是相当常见的。菱鳐属Rhombodus一直坚持至始新世时期 , Rhombodus kirklandi就是菱鳐属最后的属种了。
牛鼻鲼科(Rhinopteridae)仅只有牛鼻鲼属Rhinoptera及已灭绝的Apocopodon , Apocopodon sericeus是生存于古新世的巴西 , 于古新世后并没有化石记录 ; 牛鼻鲼属Rhinoptera于世界各地的海洋都有分布 , 牠们较偏好浅海混浊底部 , 以软体动物及甲壳类为食 。牛鼻鲼属Rhinoptera的口鼻部非常独特, 是呈二裂的 , 有菱形的背腹及长而苗条的尾部 , 骨盆位置长有一个背鳍 , 尾的底部有较多刺脊 , 牛鼻鲼属Rhinoptera的皮肤较其它鲼形目(Myliobatiformes)成员来得光滑。牛鼻鲼属Rhinoptera中以大西洋牛鼻鲼(Rhinoptera bonasus)体型最大 , 大西洋牛鼻鲼(Rhinoptera bonasus)雄鱼鳍宽79 cm , 雌鱼鳍宽61 cm , 大西洋牛鼻鲼(Rhinoptera bonasus)鳍宽最大记录为213 cm , 而大西洋牛鼻鲼(Rhinoptera bonasus)的敌人包括海鲡(Rachycentron canadum) , 高鳍真鲨Carcharhinus plumbeus及公牛鲨。
牛鼻鲼属(Rhinoptera)于古新世时期分布于北非 , 西非及欧洲 , 现在则分布于全球热带海洋。已灭绝的牛鼻鲼属种有古新世巴西的Rhinoptera prisca, 古新世至中新世的Rhinoptera raeburni , 始新世路德阶尼日尼亚及摩洛哥的Rhinoptera sherboni , 始新世伊普雷斯阶(Ypresian)至早中新世北美及非洲的Rhinoptera daviesi , 中新世早期瑞士及法国的Rhinoptera studeri , 中新世至上新世北美及非洲的Rhinoptera dubia 。于始新世伊普雷斯阶(Ypresian)牛鼻鲼属于北大西洋西部显得非常普遍 , Rhinoptera sherboni于北美维吉尼亚州也有化石发现 。
Rhinoptera raeburni是一个长寿种 , 过去只于尼日尼亚及扎伊尔的古新世地层发现牠的化石 , 不过于印度中新世地层也发现了Rhinoptera raeburni的牙齿化石。现存的牛鼻鲼属种约于渐新世至中新世出现 , 于北美卡尔弗特渐新世至中新世地层发现了大西洋牛鼻鲼亲近种(Rhinoptera aff bonasus)及巴西牛鼻鲼亲近种(Rhinoptera aff brasiliensis)的化石 。
鲼科(Myliobatidae)源于晚白垩纪时期 ,已灭绝的鲼科成员包括森诺曼阶北美的Enantiobatis , 坎帕阶至马斯特里赫特阶北美的短根鳐属Brachyrhizodus , 马斯特里赫特阶印度 , 西班牙及非洲的Igdabatis , 马斯特里赫特阶摩洛哥的Ixobatis , 古新世摩洛哥的Garabatis , 始新世乌兹别克的Aktaua , 始新世摩洛哥的伪鹞鲼属Pseudoaetobatus, Leidybatus , Lophobatis 及Zygzabatis ,始新世西欧的原鲼属Promyliobatis , 渐新世德国的Weissobatis 及上新世北美的Mesibatis。短根鳐属种Brachyrhizodus wichitaensis于晚白垩纪的北美海洋是很常见的 , 牠的牙齿保留了牛鼻鲼属的特征 , Brachyrhizodus wichitaensis被视作是原始的鲼科成员。
鲼属分布于环球温带及热带海洋 ,于海湾 , 河口的底部常可看到鲼属 , 牠们以虾类 , 蟹类 , 双壳类(Bivalvia) , 腹足类 (Gastropods) , 多毛类动物(polychaete)及鱼类为食 , 牠们会借胸鳍挖出泥沙底部的食物。 鲼属多达12种 , 其中东太平洋的加州鲼(Myliobatis californicus)及东大西洋的鲼(Myliobatis aquila)体宽可达1.8米 , 加州鲼(Myliobatis californicus)重量则可达91公斤 , 而澳大利亚鲼(Myliobatis australis)体宽可达1.6米 , 全长(包括尾部)可达2.4米, 重量至少达87公斤。至于生活于西大西洋海湾的弗氏鲼(Myliobatis freminvillii) , 西大西洋及加勒比海的古氏鲼(Myliobatis goodei)体型较细小 , 体宽少于1米。
鲼属(Myliobatis)是相当古老的鳐类 , 要追溯至古新世时期 , 科佩亚鲼Myliobatis copeanus就是被发现于马里兰州古新世Aquia组 , 此外于非洲古新世至早始新世地层发现了强齿鲼Myliobatis sulcidens及思扎里鲼Myliobatis nzadinensis 的牙齿化石; 其它鲼属化石于始新世地层都有化石记录 , 如摩洛哥的条纹鲼Myliobatis striatus及狄克森鲼Myliobatis dixoni , 南卡罗莱纳州的Myliobatis toliapicus及北卡罗莱纳州的宽齿鲼Myliobatis latidens。于欧洲渐新世地层及美国中新世地层都发现了鲼属的牙齿化石 , 渐新鲼Myliobatis oligocaena 就是生活于渐新世比利时的鲼属 , 锯齿鲼Myliobatis serratus则是渐新世德国的鲼属。至于美国马里兰州中新世地层发现的巨鲼Myliobatis gigas (Myliobatis vicomicanus)牙齿是发现众多史前鲼属牙齿中最大的 , 最大的中央牙齿宽度能达8厘米。
现存的鲼属成员只有加州鲼(Myliobatis californicus)于圣地亚哥更新世地层留下化石记录。
前鳍鲼属(Pteromylaeus)是生活于热带及温带海洋的中小型鲼科成员 , 于海湾常可看见其踪影。Compagno于1999年确认了该属有两种 , 横纹前鳍鲼(Pteromylaeus bovinus)是生活于大西洋东部(葡萄牙至南非), 地中海及印度洋西南部 , 体宽达1.8米 ; 糙前鳍鲼(Pteromylaeus asperrimus)于中太平洋东部(巴拿马及加拉帕戈斯群岛)的热带水域 , 体宽达0.8米。现存的横纹前鳍鲼(Pteromylaeus bovinus)于南黎巴嫩及法国中新世地层 , 委内瑞拉早上新世地层有化石记录 , 于中新世大西洋西部及加勒比海也存有另一已灭绝的属种Pteromylaeus apenninus。
鹞鲼属(Aetobatus)是生活于所有热带及亚热带水域沿岸的鲼科成员 , 牠一共有四种 : 纳氏鹞鲼(Aetobatus narinari)于环球热带及亚热带浅岸 , 于珊瑚礁很常见 , 牠能长达3米及重达200公斤 ; 无斑鹞鲼(Aetobatus flagellum)是生活于印度洋及西太平洋 , 属大洋底栖性 , 但于近海岸及热带海洋也有栖息 , 体宽约72厘米 ; 睛斑鹞鲼(Aetobatus ocellatus)于印度 , 爪哇及印度尼西亚一带的印度洋都有报导 ; 第四种网纹鹞鲼(Aetobatus reticulatus)的数据则非常稀少; 尽管鹞鲼属(Aetobatus)的外型和其它鲼科成员非常相似 , 但牠们有较厚的身躯 。牠们会季节性聚集 , 像纳氏鹞鲼(Aetobatus narinari)会用吻部把软体动物等挖起进食 , 牠们也进食蠕虫 , 甲壳类 , 头足类及鱼类。
最古老的鹞鲼属(Aetobatus)是古新世至始新世的非洲 , 欧洲及马里兰州的Aetobatus irregularis , 另一种较有名的已灭绝成员是发现于欧洲及英国大西洋海岸盆地中新世地层的Aetobatus arcuatus, 其它史前鹞鲼属种有始新世西欧的Aetobatus sulcatus , 中新世英国的Aetobatus rectis 及中新世北美的Aetobatus smithii。至于纳氏鹞鲼(Aetobatus narinari) , 于英国更新世地层才有化石记录。
近来于古巴中新世中期地层发现的波氏鹞鲼Aetobatus poeyi是属于体型巨大的鹞鲼属种 , 牠留下了齿板及尾刺的化石 , 有报导指波氏鹞鲼Aetobatus poeyi体型能赶上双吻前口蝠鲼(Manta birostris) , 至于这报导是否属实还需要以后论文发布来支持。
无刺鲼属(Aetomylaeus)是生活于印度洋及西太平洋大陆架及岛架 , 该属没有尾刺 , 牠们可能是半大洋性 , 一共有四种 :
花点无刺鲼(Aetomylaeus maculatus)是生活于沿岸近海的属种 , 于印度 , 印度尼西亚 , 马来西亚及中国都可见其踪影 , 鳍宽可达200cm。
鹰状无刺鲼(Aetomylaeus milvus)同时是大洋底栖性及沿岸近海的属种 , 从红海至中国 , 南至印度尼西亚都有分布。
聂氏无刺鲼(Aetomylaeus nichofii)于沿岸及远洋都有广泛的分布 , 栖息深度至少达70米 , 从波斯湾至菲律宾 , 北至韩国 , 南至澳洲都有分布 , 牠的体型细小 , 鳍宽只有58 cm。
蝠状无刺鲼(Aetomylaeus vespretilio)是中型属种 , 鳍宽达160 cm , 牠是大洋底栖性属种 , 从莫桑比克至西太平洋都有分布 。蝠状无刺鲼于珊瑚礁也很常见 , 栖息深度可达110 m。
无刺鲼属(Aetomylaeus)于北美早始新世地层已有化石记录 , 于北美及欧洲中新世至上新世地层也有化石记录 , 可惜的是化石全没被定种。
中新世早期至中期北美的无刺鲼属种(Aetomylaeus sp)化石
蝠鲼科(Mobulidae)可能源于晚白垩纪时期 , 于美国科罗拉多 , 堪萨斯州及南达科州 , 加拿大萨斯喀彻温省森诺曼阶至土伦阶地层上发现了一种奇特的软骨鱼类白垩前口蝠鲼Cretomanta canadensis的牙齿化石 , 牙齿化石仅只有1.5毫米长 , 由于牙齿实在太细小 , 加上牙齿的独特 , 因此有专家认为白垩前口蝠鲼Cretomanta canadensis和蝠鲼科或者和鲸鲨科有关系 , 白垩前口蝠鲼Cretomanta canadensis可能只是一种纯滤食的软骨鱼类 , 但归属上仍存有疑问的。
第三纪蝠鲼科(Mobulidae)于欧洲 , 北美 , 非洲及亚洲都有化石记录 , 已灭绝的蝠鲼科(Mobulidae)属系包括古新世至始新世中期的古前口蝠鲼属Archaeomanta , 晚古新世至始新世中期的Burnhamia , 始新世中期至晚始新世的始蝠鲼属Eomobula , 晚始新世的Eoplinthicus , 中新世的副蝠鲼属Paramobula 及中新世早期至中期的Plinthicus 。于众多已灭绝的第三纪蝠鲼科中 , 最值得详说的是Plinthicus , Plinthicus属一共有两种包括渐新世比利时的Plinthicus kruibekensis及中新世北美及法国的Plinthicus stenodon 。Plinthicus属的牙齿形态类似现存的蝠鲼属(Mobula) , 从牠们的牙齿来看 , Plinthicus属以软体动物为食 , 牠们是属于大洋性鳐类 。 从渐新世比利时的Plinthicus kruibekensis牙齿化石来看 , 牠们的齿板及牙齿形状都和牛鼻鲼属(Rhinoptera)的相似 , 过去有专家假定蝠鲼科(Mobulidae)是源于牛鼻鲼科 , 不过牛鼻鲼科的化石记录要比白垩前口蝠鲼Cretomanta canadensis来得晚。
A : Archaeomanta cf. melenhorsti
B : Coupatezia woutersi
C : Pseudoaetobatus sp.
蝠鲼属是生活于热带及亚热带的鲼类 , 其中下口蝠鲼(Mobula hypostoma)于大西洋东部及北大西洋西部都有分布 , 姬蝠鲼(Mobula mobular) , 罗切氏蝠鲼(Mobula rochebrunei) , 印度蝠鲼(Mobula thurstoni) , 短翅蝠鲼(Mobula kuhlii) , 褐背蝠鲼(又称台湾蝠魟Mobula tarapacana)都是生活于东大西洋的蝠鲼属种 。牠们只有下颌是具钉状的细小牙齿 , 牙根大多是二裂的 , 牙齿并不是用作碾碎食物 , 牠们会于水表层吞食大洋性小型鱼类及浮游生物。最大的蝠鲼属种是姬蝠鲼(Mobula mobular) , 体宽能达5.2米。蝠鲼属于渐新世早期已存在 , 已知的中新世属种有二种包括中新世欧洲和北卡罗莱纳州的Mobula loupianensis及中新世法国的蓖齿蝠鲼Mobula pectinata。
前口蝠鲼属(Manta) 是滤食性鱼类 ,分布于环球热带海洋 , 牠一共有三种包括双吻前口蝠鲼(Manta birostris) , 埃氏前口蝠鲼(Manta ehrenbergii)及Manta raya , 这三种非常相似 , 后二者可能只是独立种群 。双吻前口蝠鲼(Manta birostris)有小量性双态性 , 雄鱼体宽5.2米至6.1米 , 雌鱼体宽5.5米至6.8米 , 最大个体体宽9.1米及重达3吨。
前口蝠鲼属(Manta)于早中新世已存在 , 已灭绝的属种包括早中新世北卡罗莱纳州的梅兰雅蝠鲼Manta melanyae及海恩蝠鲼Manta hynei , 海恩蝠鲼Manta hynei牙齿高度只有3 mm , 牠的齿冠及牙根和双吻前口蝠鲼(Manta birostris)的相似 , 于北卡罗莱纳州上新世地层也有未定种的蝠鲼属(Manta sp)化石发现 。
电鳐目分布在热带和亚热带近海,一般体形较小 , 卵胎生 。电鳐目鱼腮裂和口都在腹位,有五个腮裂,身体平扁呈卵圆形,吻不突出,臀鳍消失,尾鳍很小,胸鳍宽大,胸鳍前缘和体侧相连接。电鳐目下分二科包括电鳐科(Torpedinidae)及双鳍电鳐科(Narkidae) , 双鳍电鳐科下分九属包括深海电鳐属(Benthobatis), 坚皮单鳍电鳐属(Crassinarke), 双电鳐属(Diplobatis) , 盘臀电鳐属(Discopyge) , 异双鳍电鳐属(Heteronarce) , 双鳍电鳐属(Narcine) , 单鳍电鳐属(Narke) , 缺鳍电鳐属(Temera)及盲电鳐属(Typhlonarke) ; 电鳐科(Torpedinidae)只有两属包括澳洲睡电鳐属(Hypnos)及电鳐属(Torpedo) 。
电鳐类的头部两边都各有一个发电器 , 当大脑神经受刺激时 , 两个发电器便会把神经能转变成电能释出电力了 。其中体型最大的地中海电鲼(Torpedo nobiliana)全长能达1.8米及重达90公斤 , 牠释出的电能可达220伏特 。电鳐类是集高科技的鱼类 , 不过这「高科技」恐怕于早第三纪已存在了。最古老的电鳐科(Torpedinidae)成员是古新世至始新世北非及欧洲的始电鳐属Eotorpedo , 异电鳐属Heterotorpedo 及始新世欧洲的Narcopterus。此外电鳐属于始新世时期已存在 , Torpedo dormaalensis就是生存于始新世塔内提阶(Thanetian)至伊普雷斯阶(Ypresian ), 至于现存的普通电鳐Torpedo torpedo , 于法国南部上新世地层已有化石记录 , 于法国及比利时中新世地层也发现了未定种的电鳐属(Torpedo sp)化石; 双鳍电鳐科(Narkidae)的化石记录要追遡至始新世 , 莫里尼双鳍电鳐(Narcine molini)就是生活于伊普雷斯阶(Ypresian)意大利的双鳍电鳐属种。
软骨鱼类的演化史在这里也要告一段落了 , 牠们于沧海中遗下的每一颗牙齿 , 都在细说万丈光芒的历史 。回味软骨鱼类的演化史,从志留纪至第四纪一共四亿二千万年的岁月 , 牠们就像细小顽强、偶有惊艳的金鳞 , 于地球各大时代之水中游弋 , 无论那水湍急还是和缓,充沛还是涓流 , 牠们还是存在于那水中 。金鳞岂是池中物 , 遇上风云便能呼风唤雨。从板足鲎类至鲸类等各时代存在的众多对手 , 泰半已如梦幻泡沫般地消失不见 , 只有软骨鱼类还能如涛涛江水永不休 。
众多鳐目成员的生存时期 :
RAJIFORMES
?Rajiformes inc. fam. Pliensbachian (unnamed tooth [Curtis
and Padian 1999]) to Toarcian (several taxa [see Delsate and
Candoni 2001]).
‘‘Spathobatidae.’’ Toarcian (Belemnobatis moorbergensis [Thies
1983]); to Aptian (Belemnobatis picteti [Cappetta 1970] or
Maastrichtian if Squatirhina belongs within this clade (Underwood
et al. 1999).
Rajiformes inc. fam. A number of taxa are known from isolated
teeth that cannot be readily accommodated within other families.
Examples exist within the Kimmeridgian (Engaibatis
schultzei Arratia et al. 2002; Leiribatos alienus Kriwet 2004) and
throughout the Cretaceous (including such genera as Pseudohypolophus
and Turoniabatis).
Rhinobatidae s. l. Tithonian (unidentified skeletal remains
[Cione 1999]) to Recent.
Rhynchobatidae. Maastrichtian to Recent. Possibly present in
the Santonian (personal observation).
Rajidae. Cenomanian to Recent.
Platyrhinidae. Campanian/Maastrichtian (Tethybatis selachoides
Carvalho 2004) to Recent.
Cyclobatidae. Cenomanian.
Hypsobatidae. Campanian to Maastrichtian.
Parapalaeobatidae. Santonian to Maastrichtian.
Dasyatidae. Hauterivian (Dasyatis speetonensis Underwood et al.
1999) to Recent.
Gymnuridae. Cenomanian to Recent.
Myliobatidae. Maastrichtian to Recent.
Rhombodontidae. Maastrichtian.
Sclerorhynchoidei. The taxonomy of this group requires more
study, and it is likely that more than one family is present.
Barremian (Onchopristis dunklei McNulty and Slaughter 1962
[Kriwet 1999]) to Maastrichtian.
最后交代一下波氏鹞鲼Aetobatus poeyi的数据出处,是否可靠不得而知
波氏鹞鲼Aetobatus poeyi说不定是有史以来最大的鳐类!
Esbozo de una Aetobatus poeyi, raya águila prehistorica que vivio durante el Mioceno y segun se dice, alcanzaba los diez metros de envergadura. No es tan exagerado como parece; la manta actual alcanza los 8.5 metros.
Como todas las rayas águila, es muy posible que esta careciera de aguijón, pero tenia unas mandibulas muy poderosas. Hay quienes la llaman "un rival del Megalodon", pero su estilo de vida era muy diferente.
Hubo mas de una especie gigante de Aetobatus en epocas prehistoricas, y al parecer tambien habia rayas gigantes del genero Raja.
Sketch of a giant Aetobatus poeyi, a prehistoric eagle ray that lived during the Miocene and could have a span of 10 meters. It isn´t as far fetched as one could think; today's manta rays can reach 8.5 meters. As all other eagle rays, it is likely that this one lacked a stinging barb, but instead had very powerful jaws. It has been called "a rival to Megalodon", but its lifestyle was very different.
There were several species of giant Aetobatus in prehistoric times, and it seems that there were also giant skates (genus Raja).
(本文为化石网论坛精华帖,由网友景德泰岚提供,仅供大家学习参考。编辑:常君滢)