



美人鱼 , 这名字相信大家也不会陌生了 , 不论于中国 , 日本及西方诸国都会有所流传 , 欧洲传说中人鱼上半身是女性 ,下半身是鱼的身体,伴随着长长的秀发漫游特提斯海。所谓的人鱼 , 其实就是现存的儒艮及海牛 , 当时的人们很可能看错了,把这些动物看成怪物并加以流传。而这些所谓的美人鱼 , 除了有段名垂千古的传说外 , 背后亦曾有过一段光辉岁月, 以下为本人根据一些连结而整理的资料 , 从而揭开特提斯美人的神秘面纱 !
海牛目跟长鼻目Proboscidea 关系较接近 , Tethytheria 是由海牛目(Sirenia) , 长鼻目(Proboscidea) , 绝种的索齿兽目(Desmostylia)及重脚目(Embrithopoda)组成 。特提斯兽Tethyeria看似是演化出最原始的有蹄哺乳类髁节目 (condylarths) , 出现于特提斯海(古地中海)的海岸 。
根据形态学及分子学研究指出 , 海牛目的祖先跟鲸豚类及鳍脚类等祖先关系遥远 , 尽管大家都是同时于水中生活及进化。海牛目祖先于古新世(6500万年至5400万年前)跟其近亲分道扬镳并迅速重返水中 , 后更扩展至全世界。
海牛目最初进化时 , 它们本身已是素食者 , 依靠海草及水生的被子植物为食。直至现在 , 它们是生活于热带海洋并仍然以被子植物为食 。海牛目拥有较大的肺部 , 沉重的骨骼以助它们于浅水中作停留 。 它们的骨骼是肿胀 , 密集脓厚的 , 特别是肋骨 , 经常可于化石中找到。始新世的海牛目成员 , 跟中生代的哺乳类相似 , 相反跟其它新生代的形成对比 , 前臼齿是有5颗而非4颗 , 齿式是3.1.5.3 。现存的海牛目成员则大多保留了一对獠牙似的上颚门齿 , 前方的牙齿如门齿及犬齿都丧失 , 跟早期的齿式不同。
于印度西北部早始新世的Subathu组发现的Ishatherium subathuensis仅只有臼齿M2化石发现 , 化石有四个主齿尖及呈圆锥形 , Ishatherium subathuensis是否属于海牛目是存有疑问的。此外一些海牛目成员如始新世前期的Paralitherium tarkanyense , 始新世中期匈牙利的Sirenavus hungaricus , 始新世中期欧洲的Anisosiren pannonica , 渐新世哥伦比亚的Lophiodolodus chaparralensis , 中新世斯里兰卡的Miodugong brevicranius及中新世法国的Prohalicore dubaleni的分类地位都存有疑问 , 且缺少具体资料 , 在这贴中不详细描述了。
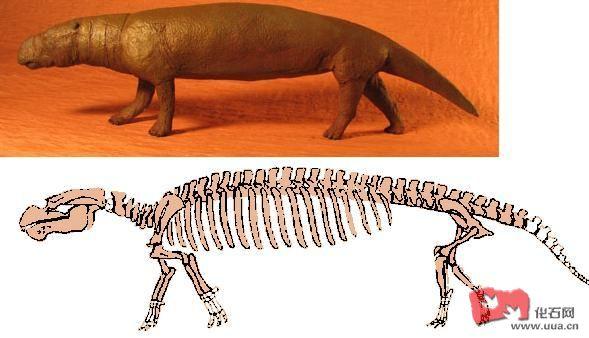
Prorastomidae科 (始新世中期)留有最古老的海牛目化石纪录 , 是类似古新世至始新世髁节目的两栖四足动物 , 但却变得水生专门化 , 例如有收缩的鼻腔 , 鼻侧的气窦丧失 , 肋骨变得肿胀及密集。最原始的Prorastomids 属种能透过骶骨间的关节支持身体重量以便于陆地上移动。头骨前方及下鄂骨显得不偏斜及横向扁平 , 后肢有较为良好的发展。Prorastomids 拥有较直的喙 , 然而后来的海牛目成员不多不少拥有朝下方的猪鼻状突出物。
Prorastomus 结构上介乎特提斯兽tethytheres 及后期的海牛目成员之间 , 尽管Prorastomus可能不是后期所有海牛目成员的直系祖先 , 但是Prorastomus已明显具备一些已知的海牛目成员特征了 , 包括膨大的喙 , 吻突 , 收缩且扩大的鼻孔及五颗前臼齿 。
Prorastomus sirenoides 生存于始新世中期的牙买加 , 骶骨间的关节缩小 , 但仍能于陆地上承托其身体重量。
Prorastomidae中还有一个未命名的种被发现于牙买加的Chapelton组。
Pezosiren portelli (牙买加) 拥有最古老的海牛目化石纪录 , 这种动物有良好发育的四肢 , 骶骨由多椎骨组成并有较强的骶髂关节连接从而于陆地上支撑其身体重量 , Pezosiren portelli 很可能经常于水中生活 。 Pezosiren portelli 以后的中间种化石显示进化的转变是介乎陆栖及水栖之间 , 海牛目的进化可能类似同时代的原始鲸类 , 透过脊椎延长及骨盆位置同时成了浆状物 , 后肢丧失及尾部变大以作划水游动。
原海牛科Protosirenidae
原海牛科Protosirenidae ,生存于始新世中期 , 第二类重要的早期海牛目成员 ,虽然仍保留了良好发展且短小的后肢 , 但肢间关节不牢固 , 这正可说明了于陆地上其后肢是难以支撑其身体重量。原海牛科Protosirenidae中的原海牛属Protosiren于始新世中期得以大量发展扩散 , 由北非的埃及至南亚的印巴间的特提斯海(古地中海)皆有其分布。透过其头骨及部份骨骼 , 得以知道原海牛属Protosiren 是主要生活于海中并甚少留在陆地上的动物。原海牛属Protosiren除了拥有发展良好且短小的后肢外, 口部还有朝下方的猪鼻状突出物以便于海底进食。下颌联合变宽意味着其植食的选择性缩小 , 其犬齿及门牙缩小并最终丧失(不包括獠牙) 。Prorastomid 及原海牛属Protosirenids 最终于始新世晚期(3800万年前) 灭绝。
原海牛属Protosiren 于海牛目中是与别不同的在于胸椎横切面上拥有较大的钥匙孔形的神经中枢管道 , 肋骨首端一般是软骨而非滑膜关节的 , 并且肋骨是缺乏骨肥厚(pachyostosis)。Protosiren eothene 跟其它原海牛属品种不同之处在于体型细小 , 前胸中枢较Protosiren fraasi的要短10-12% 。 Protosiren eothene椎骨关节部分的肋骨首端至少是滑膜的 , 并拥有良好但相对较肋骨首端细小的肋骨结节。此外 Protosiren eothene颈椎及子宫颈椎骨较后期的原海牛属Protosiren 成员来得长 , Protosiren eothene颅基长可达350mm 。
原海牛属Protosiren (埃及 , 北美 , 欧洲及亚洲 )
1. Protosiren eothene , 生存于早始新世中期的巴基斯坦 。
2. Protosiren fraasi , 生存于早始新世中期的印度 , 埃及及匈牙利。
3. Protosiren sattaensis , 生存于晚始新世中期(3900万年前至4000万年前) 的巴基斯坦。
4. Protosiren smithae , 生存于晚始新世中期的埃及 , 为Protosiren fraasi 直属后代 。
5. Protosiren minima , 生存于始新世中期至晚期的法国。
原海牛科Protosirenidae也有新属种于北卡罗莱纳州的Avon Park , Inglis 及Castle Hayne组有所发现 , 另外有些属种化石可能来自佛罗里达晚始新世的Crystal River组 。
原海牛属Protosiren开始发展出类似现今海牛科成员的骨骼 , 但齿式仍然原始落后 , 被认为分为两个不同的世系 :
儒艮类 : Eotheroides (中始新世)拥有较为轻微弯曲的猪状形吻部及细小的獠牙 , 同时也拥有原始的齿式 。后来于渐新世时期兴起了Halitherium 而这些儒艮科成员拥有较弯曲的猪状吻部及较长的獠牙 。当然现今的儒艮类的吻部更为弯曲 , 獠牙更大。
海牛类 : Sirenotherium (早中新世) , Potamosiren (晚中新世 ) 等类似现今海牛类成员已丧失了部分颊齿 ; 早上新世时期 Ribodon 牙齿则具连续替换的能力。
儒艮科Dugongidae
儒艮科从始新世中期/晚期一直延存至现在 , 儒艮科早期的成员已是完全水生 , 并保留退化的后肢 , 这是相当成功的海牛家族类群并于后来兴起了海牛科 , 渐新世时期(3800万年前至2300万年前)一度是儒艮科的黄金时期 。儒艮科下分Halitheriinae亚科及儒艮亚科Dugonginae , Halitheriinae亚科包括Eotheroides , Prototherium ,始海牛属Eosiren , Halitherium, Metaxytherium 及加勒比海海牛属Caribosiren , 儒艮亚科Dugonginae包括Dioplotherium , Corystosiren 及Rytiodus等于中新世早期均有化石纪录 。
于中新世后(结束于500万年前) , Halitheriinae亚科在加勒比海便消声匿迹 , Halitheriinae亚科有以下属种:
Eotheroides : Eotheroides aegyptiacum 化石被发现于埃及开罗的始新世中期地层 , 稍后于始新世晚期的埃及法扬(Fayum)海床也有所发现 , 于2006年印度发现的新种Eotheroides babiae仅只有下颌及牙齿化石发现。始新世中期北非的Eotheroides属被视作最古老的儒艮科成员 , 前上颌骨有一对细小的獠牙 , Eotheroides属有双脊牙型的臼齿 。Eotheroides属和晚始新世南欧的Prototherium属有密切的关系 , 二者都保留了3.1.5.3 的齿式 。
Prototherium : Prototherium veronense , 生活于始新世晚期的意大利 , 最原始的儒艮科成员之一 , 未成年的齿系相当清楚 , 拥有5颗原始的前臼齿 。过去一直没有未成年的儒艮科齿系化石的发现 , 但是Prototherium veronense 的齿系发现却令人震惊 , 因齿系跟现存的海牛属(Trichechus)的相似 。 此外Prototherium还有Prototherium intermedium被发现。
始海牛属Eosiren , 生存于始新世中期至晚期的地中海 , 有以下属种 :
1.Eosiren abeli , 生存于渐新世早期的埃及
2.Eosiren libyca, 晚始新世
3.Eosiren stromeri
4. Eosiren imenti , 早渐新世的埃及
始海牛属Eosiren大多只有下颌化石 , 众多始海牛属种以伊曼地始海牛Eosiren imenti于海牛目进化史中占最重要的角色。伊曼地始海牛Eosiren imenti的颅基可长达400mm , 从伊曼地始海牛Eosiren imenti发现的地点来看 , 伊曼地始海牛Eosiren imenti有很高的耐盐性 。伊曼地始海牛Eosiren imenti 和晚始新世的Eosiren libyca 及 "Prototherium" intermedium相似 , 于系统分枝学上和Halitherium schinzii 有密切关系 , 伊曼地始海牛Eosiren imenti可能是源于Eosiren libyca 。
伊曼地始海牛Eosiren imenti被认为可能和后来的儒艮亚科Dugonginae,中新海牛亚科Miosireninae及海牛亚科Trichechinae的起源有关 , 儒艮亚科Dugonginae首次于晚渐新世美国东南部出现 , 已知最古老的儒艮亚科属种是Crenatosiren okieni , 此外最古老的海牛科则是晚渐新世的Anomotherium 。
Halitherium出现于晚始新世时期并延存至晚渐新世时期的欧洲 , 并显然于大西洋及加勒比海演化成Metaxytherium 。Halitherium鳍状肢中的趾骨间存有间隙, 后肢退化但趾骨已不存在了 , 不过Halitherium还存有基本的大腿骨连接着退化的骨盆上 , Halitherium 有延长的肋骨增加肺容量以便有效控制浮力 。Halitherium taulannense被发现于2001年 , 生存于晚始新世的法国 , Halitherium taulannense的形态介乎于始新世的海牛目成员及早渐新世欧洲的Halitherium schinzii。
Halitherium属种如下:
1.Halitherium schinzii
2.Halitherium christolii
3.Halitherium antillense
4.Halitherium alleni
5.Halitherium taulannense
加勒比海海牛属Caribosiren只有Caribosiren turneri一种 ,生存于晚渐新世时期的波多黎各(Puerto Rico)。加勒比海海牛属Caribosiren头骨可达300 mm, 吻部偏斜75度及没有獠牙 , 可能专门以海草及根茎为食 。于渐新世结束后再没有加勒比海海牛属Caribosiren的化石记录。
Metaxytherium生存于中新世至上新世 , 跟Crenatosiren及Dioplotherium共存于北佛罗里达 , 跟杜氏海牛属Dusisiren 及 Dioplotherium 共存于墨西哥的下加利福尼亚州 , 分类学上Hesperosiren 跟 Metaxytherium 同义 。Metaxytherium属种有明显向下导向的吻部 , 有细小至中度大小的獠牙 , 以长在水深超过1米的细小海草为食 。
Metaxytherium krahuletzi , 生存于早期中新世的欧洲 ,可能是所有Metaxytherium 成员的直系祖先。
Metaxytherium medium , 生存于欧洲 , Metaxytherium krahuletzi的直系后代。
Metaxytherium serresii , 生存于欧洲 , Metaxytherium medium的直系后代。
于上新世时期 , 其中部分海牛离开地中海 , 它们代表着欧洲至北非间特提斯海中最后的海牛并灭绝于上新世晚期 。于北非利比亚中的Sahabi组中 , 于Metaxytherium serresii 骨骼中发现了一些咬痕 , 显示可能受大型鲨鱼攻击 ,并于Metaxytherium serresii 化石地四周找到巨齿鲨(Carcharodon megalodon)化石 , 意味着当时鲨鱼会捕食海牛目并扮演着重要的生态角色。
Metaxytherium subapenninum(Metaxytherium forestii ) , 生存于欧洲, Metaxytherium serresii的直系后代。
Metaxytherium crataegense(Metaxytherium calvertense/ Metaxytherium riveroi) 全长约3米 ,生存于中新世中期 , 散布于美洲大西洋 , 东太平洋墨西哥的下加利福尼亚州及加利褔尼亚洲一带。
Metaxytherium crataegense 的化石骨架,拥有1500万年之久的历史,于1929年,被发现于佛罗里达州西北部 ,现收藏于佛罗里达州国立大学的地质学研究室。
Metaxytherium arctodites ,分布于旧金山一带 ,跟无齿海牛亚科Hydrodamalinae 有直属关系。Metaxytherium arctodites颅基长可达415mm。
儒艮亚科Dugonginae是儒艮科中唯一生活于上新世时期的加勒比海一带 , 于这时期加勒比海一带的Halitheriinae化石纪录亦消失不见 , 异海牛属Xenosiren(Dioplotherium后代)可能跟Corystosiren一样存活于上新世早期。 晚上新世时期加勒比海地区有一种类似现今儒艮的儒艮科成员 , 但却拥有较大的獠牙(未命名), 它的直系祖先不明 。于晚上新世时期(180万年前) , 儒艮类看似于西大西洋全部灭绝 。 儒艮亚科有演化出较大及刀片状的獠牙趋势 , 对比渐新世至中新世时期Halitheriine那些细小且近似圆锥的獠牙 , 儒艮亚科Dugonginae 更能有效进食海草的根茎。儒艮亚科繁衍于加勒比海至西大西洋地区及进行幅射形进化 , 于晚渐新世至早中新世时期分布于全世界的热带地区 。
Crenatosiren olseni生存于渐新世晚期的美国东南部 , 相比后来的儒艮亚科成员 , Crenatosiren的獠牙比Metaxytherium的要来得大 , 被认为是用作挖掘根茎 。
儒艮(Dugong dugon ) 这是唯一现存的儒艮亚科成员 , 现分布于三十七个国家 , 由非洲西南方和马达拉斯加岛四处的印度洋海岸线 , 包括红海 , 马纳尔湾(印度半岛南端及斯里兰卡之间) , 鲨鱼湾 , 西澳大利亚 , 印太平洋群岛 , 托雷斯海峡(Torres Strait) , 太平洋则南达赫维湾(Hervey)及摩湾(Moreton) , 北达日本冲绳岛。
儒艮的体型大而呈纺锤状,体长约2.4至2.7米,3米以上的个体相当少见,一般而言雌性的体型会比雄性大一点 。皮肤光滑,外观呈褐至暗灰色,腹部颜色较背部来得浅,体表毛发稀疏。颈部短,但仍能有限度的转动头部或点头。前肢短、呈鳍状,末端略圆而缺乏趾甲;胸鳍是幼儒艮主要的推进力来源,成年后则转变为以尾鳍为主。乳房1对,乳头位于前肢基部处。儒艮没有外耳壳,只看得到小小的耳孔,眼睛也很小。鼻孔位于吻部顶端,周围有皮膜可在潜水时盖住鼻孔。宽而扁平的嘴位于厚重吻部的末端下方,嘴边的短须是进食时的重要工具。儒艮有2对门齿,上、下颚各有3对前臼齿与3对臼齿,但所有牙齿不会同时存在,随着年龄增长,它们会失去第1对门齿、所有的前臼齿与第1对臼齿,剩余的两对臼齿则会终生成长。雄性的第2对门齿在12-15岁时会穿透上颚外露,但当闭上嘴时会被厚重的上唇盖住而看不见;雌性极少见到外露的门齿,但少数会保留至30岁左右。
儒艮的肺很大,从胸部一直延伸至肾脏附近,由呈水平的横隔膜将其与其它脏器分隔。支气管深入肺的大部分区域,细部分支少且大半由软骨构成,目前仍不清楚儒艮潜水时肺部是否会和鲸类一般有塌陷的情形。跟其它海洋哺乳动物比起来,它们的脂肪层厚度较薄,同时身体周边似乎不存在热逆流交换系统(heat countercurrent exchange system),肢体末端的动脉与静脉紧邻以维持体热不散失的一种血管构造),这可能与它们栖息于温暖海域有关。胃的构造简单,大肠很大(达胃的两倍重),推测是纤维素的主要消化场所,长度达25米以上,相当于小肠两倍的长度。
一些儒艮亚科成员如Rytiodus, Corystosiren, 异海牛属Xenosiren及Dioplotherium等属有扩大的刀片状獠牙以掘取海草 , Rytiodus及Corystosiren的獠牙比Dioplotherium还要宽阔及更锐利。现存的儒艮也有演化出较大的獠牙 , 但看似只是用作社交。
Dioplotherium属
Dioplotherium manigaulti 分布于晚渐新世的佛罗里达 , 属较分歧的专科并跟Crenatosiren 属同时期的儒艮亚科成员。 Dioplotherium manigaulti及Metaxytherium floridanum头骨均可达460 mm。
Dioplotherium allisoni 于早中新世时分布于巴西 , 加利褔尼亚和墨西哥下加里福尼亚州 ; 中新世中期则分布于阿根廷。于加利福尼亚Dioplotherium allisoni 跟史德拉大海牛的祖先杜氏海牛属Dusisiren 分布有所重叠。Dioplotherium allisoni下颌偏斜度比吻突还要大15至20度 , 牠有着Dioplotherium manigaulti及Rytiodus的特征 , Dioplotherium allisoni被认为是异海牛属Xenosiren的先祖。
异海牛属Xenosiren 仅只有尤卡坦异海牛Xenosiren yucateca , 分布于晚中新世至早上新世的尤卡坦半岛 , 为Dioplotherium allisoni的直属后裔。尤卡坦异海牛Xenosiren yucateca和Corystosiren varguezi同样有着较大的獠牙 , 尤卡坦异海牛Xenosiren yucateca有独特的头骨结构 , 下颌前端有紧密的嚼肌可推动獠牙来挖掘根茎 , 但上颌的牙槽突退化 , 被认为以较细小的根茎进食。Dioplotherium及异海牛属Xenosiren世系于上新世时期完全灭绝。
Corystosiren varguezi分布于上新世早期的尤卡坦半岛及佛罗里达 , 但世系也可来贯及中新世时期的当地 , 独特之处在于拥有非常厚重结实的头盖骨。
Rytiodus capgrandi生存于中新世 , 分布区域跟Dioplotherium allisoni及 Metaxytherium重叠于欧洲 , 北非 及巴西 。 Rytiodus capgrandi长度几乎是现今海牛类的两部 , 全长6米 , 拥有一对鳍状物 , 流线型的身体及尾鳍 。 较平坦的猪状形吻部可容许Rytiodus于较浅的海岸进食 , 不同其它儒艮成员 , Rytiodus capgrandi拥有较短的突出獠牙以榨取沙中的食物。
巴拉特海牛属Bharatisiren一共有两种包括印度巴拉特海牛Bharatisiren indica及喀奇巴拉特海牛Bharatisiren kachchhensis ,印度巴拉特海牛Bharatisiren indica属晚渐新世时期 , 牠的发现显示儒艮亚科是于晚始新世或早渐新世时期越过大西洋进入印非地区 ; 至于喀奇巴拉特海牛Bharatisiren kachchhensis属中新世 , 形态和印度巴拉特海牛Bharatisiren indica的很相似 , 喀奇巴拉特海牛Bharatisiren kachchhensis可能和现在的儒艮属起源相当接近。
于印度也发现了其它儒艮亚科的化石包括早中新世的印度海牛属种Indosiren koeningswaldi 及Domningia sodhae , Indosiren koeningswaldi只有不完整的化石 , 于印度尼西亚也发现了另一属种爪哇印度海牛Indosiren javanense , 属中新世中期 , 印度海牛属Indosiren的数据不流通。
Domningia sodhae被发现于印度喀曲 , 属早中新世 。Domningia sodhae的颅基长可达510 mm , 属体型较大的儒艮亚科成员 。 Domningia sodhae和巴拉特海牛属Bharatisiren前臼齿同样有着多牙根 , 前上颌骨有着较长的鼻突 , 二者的上腭形态都很相似 , 有平坦的剪切型的獠牙。Domningia sodhae , 巴拉特海牛属Bharatisiren及Dioplotherium manigaulti被视作儒艮亚科的基层类群。
侏儒海牛属Nanosiren属于儒艮亚科基层成员 , 一共有二种包括晚中新世委内瑞拉的桑切氏侏儒海牛Nanosiren sanchezi及早上新世佛罗里达的加西亚侏儒海牛Nanosiren garciae , 加西亚侏儒海牛Nanosiren garciae被认为源于桑切氏侏儒海牛Nanosiren sanchezi。加西亚侏儒海牛Nanosiren garciae全长仅只有2米及重达150公斤 。侏儒海牛属Nanosiren有细小的圆锥状獠牙 , 吻部偏斜 , 侏儒海牛属Nanosiren可能以沿岸的二药藻属(Halodule)及喜盐草属(Halophila)等细小的水生植物为食。
除了上述的儒艮属成员 , 还有一种未被命名的新种 , 生存于晚上新世时期的佛罗里达 , 跟现存的印太海牛关系接近 , 最近才被认知的加勒比海及西大西洋地区的儒艮亚科成员。
北太平洋的Metaxytherium 属种于中新世时期进化成无齿海牛亚科Hydrodamalinae 并以史德拉大海牛灭绝作终结 , 无齿海牛属(Hydrodamalis)中最大成员更可长达九米以上。这些种类都能成功适应气候及寒冷的水温 , 并以海藻类为食。无齿海牛属(Hydrodamalis) , 固名思义 , 就是完全没有牙齿 , 并且拥有缩短 , 脚抓似的鳍状肢用来收集植物及避开暗礁 ,同时亦没有趾骨。人类对史德拉大海牛的过渡捕杀使其灭绝于十八世纪。
无齿海牛亚科Hydrodamalinae特征在于新的适应方式 , 跟其它海牛目成员的底部觅食方法背离 , 体型增大 , 扩大领域至寒冷的水域 , 能进一步适应太平洋冰冷的水温并以大型褐藻作食粮取代原本以海草为食。
杜氏海牛属Dusisiren
Dusisiren jordani , 生活于中新世中期(1200万年前至1000万年前)的加尼褔尼亚 , 为 Dusisiren dewana 直属祖先 , 分布区域跟Metaxytherium 及 Dioplotherium 重叠 , 全长4.3米。
Dusisiren dewana , 生活于中新世中期(1000万年前至900万年前)的日本 , 形态学及年代学介乎于杜氏海牛属及无齿海牛属之间 , 全长3至4米。
无齿海牛属Hydrodamalis
Hydrodamalis cuestae是生存于上新世前期(由5百万年至3百万年前) , 分布于加利褔亚及墨西哥的下加利福尼亚州California & Baja 一带。Hydrodamalis cuestae 的头骨中亦是海牛成员中最大的 , 同时亦是有史以来最大的海牛目成员 , 长达10米, 重达22,046磅 , 亦被认为是史德拉大海牛Hydrodamalis gigas的祖先 。
日本北太平洋发现了无齿海牛属Hydrodamalis spissa的化石 , 和杜氏海牛属有一定的相似性。于Dusisiren dewana , Dusisiren takasatensis及 Hydrodamalis spissa , 它们的吻突功能上的发展和史德拉大海牛相似 , 它们颌骨咀嚼轮廓狭窄及矩形 , 翼突腹侧宽大且和翼状肌相连。 Dusisiren dewana及 Dusisiren takasatensis 的夹肌发展类似Dusisiren jordani而不像Hydrodamalis spissa , 半头夹肌的发育则类似史德拉大海牛Hydrodamalis gigas 。Dusisiren takasatensis 有其它特征 , 鳞状骨有较长的S形脊 , 有较大的枕髁及出众的枕蝶骨 , 相对Dusisiren dewana , Dusisiren takasatensis外枕骨有较厚的侧缘。Dusisiren takasatensis 有强健的夹肌 , 半棘肌及头长肌 , 颈部有足够的力量可把头部升降。于无齿海牛亚科中胸椎中部有较低的刺突 , 这意味着轴上肌的发育较少, 椎骨柱灵活性较慢 , 它们游动时像静止一样。
史德拉大海牛Hydrodamalis gigas 于19世纪时曾被归归Rytina 。于1741年在白令海峡发现,发现者为博物学者乔治.斯特拉,当时他正与探险家维他斯.白令一同旅行,而他也是已知唯一看过大海牛活体的生物学家。
史德拉大海牛首次被人类发现时,其分布范围大致位于白令海的司令群岛周边,往东达阿留申群岛西侧,往西至堪察加半岛。由上新世(Pliocene)与更新世(Pleistocene)的无齿海牛属化石记录分析, 过去史德拉大海牛的分布范围可能广达整个北太平洋中北部海岸地带,东侧达下加利福尼亚半岛与现今墨西哥西岸约北纬30度处,西部则到日本的本州岛岛岛(南限约北纬37度)。
当斯特拉第一次描述大海牛的时候,大海牛的群体已经很小,分布地区很有限,数量仅余1,000至2,000头。在人类发现它们后不久,白令海地区拥入许多猎人,这些人的目标是捕杀生存于当地的海獭与北方海狗以获取毛皮,而为了补给需求造成了大海牛的悲剧。这些人大量捕杀大海牛,吃它们的肉、用它们的皮来制造与修补船只。
部分学者认为,海獭的减少可能也是大海牛灭绝的原因之一,在其它生活于白令海的动物中,海獭可能是与它们关系最密切的哺乳动物,因为海獭会捕捉啃食海藻的海胆,如果放任海胆漫无止境的增加,很快地海藻林就会被破坏殆尽。因此当时海獭的减少可能也让大海牛目临食物短缺的压力。
最后一头大海牛纪录是于1768年被杀 , 距离被发现时整整27年便宣告灭绝 。 从史德拉形容所得 , 这巨大草食动物于北太平洋的白令岛及铜岛的数目约为 1500 至 2000 头左右 。这种巨大的动物约长约7. 5米,体重4,500至5,900公斤。 大海牛是没牙齿的 , 嘴里只有两片又平又白的骨头,一块在上颌,一块在下颌 , 两片骨头带有凸起的皱褶以便咀嚼海藻。斯特拉的记录中提到它们会“近岸集体进食,幼兽会被围在群体中央保护” ,而且似乎不会潜水,至少进食时它们只将部分身躯埋入水中,在岸边啃食潮线附近接近海面的大型藻类, 估计其觅食深度不超过1米深 , 单兽甚至可把小艇撞翻。
关于大海牛的文献记录中,曾提到有1头雌性大海牛被捕杀拖上陆地,而另1头与它同行的雄性大海牛就在岸边停留不肯离去,时间至少持续2天以上。加上其它群体观察记录,分析大海牛可能是1夫1妻制,配偶关系可能会维持相当长的时间。斯特拉留下的记录中提到,大海牛多在早春时交配,怀孕期在12个月以 上,终年可生产,不过秋天是分娩的高峰期。事实上斯特拉在白令岛停留的时间从1741年11月底至1742年的8月,由时间上看来他可能不曾亲眼目睹初生 不久的大海牛。
海牛科Trichechidae出现于晚始新世或早渐新世(约3800万年前) ,可能起源于儒艮亚科Dugongidae 。于上新世时期(500万年前至180万年前) , 海牛科Trichechidae的早期成员Ribodon 生活于北卡罗莱纳洲的加勒比海/西大西洋区域一带。于晚上新世时期 , Ribodon 把其活动范围扩展至南美之外。 Ribodon 及其后代海牛属Trichechus 有额外的臼齿 , 它们演化出特殊的牙齿替换系统,以因应食用多纤维食物造成的牙齿磨损。海牛科仅具臼齿, 更新方式不是一颗掉了后再重新长出新牙,而是整列牙齿由颚的末端水平地往前移动,当牙齿移动至颚的最前端时,牙根会逐渐被吸收终至脱落。海牛科Trichechidae可能曾被局限于南美的海河交界及沿海一带直至晚中新世才有所改变。当地它们以内河植物为食 , 而儒艮科则生活于西大西洋及加勒比海并开拓该区的海草牧场。上新世中期(1000万年前至500万年前)安第斯山脉形成时倾倒大量淤泥并把大量营养溶解于南美河流中 , 直接刺激水生的大型植物生长 , 特别是草本植物禾本科(Gramineae)。海牛科成员为了适应新的食物来源 , Ribodon首先拥有额外的臼齿能作水平替换 , 稍后臼齿的尺寸缩小 , 珐琅脊长度增加 , 珐琅褶变得复杂 , 齿排中牙齿数目增加 , 这都于海牛属Trichechus中表现出来。
中新海牛亚科Miosireninae 是海牛亚科Trichechidae 的姐妹组份 , 现已归入海牛科 , 最初海牛科Trichechidae是被认为起源于原海牛科Protosirenidae 及于始新世中期出现的, 但后来中新海牛亚科Miosireninae被认为是根源于儒艮亚科Dugongidae 并兴起于晚始新世或渐新世早期 。
Anomotherium只有Anomotherium langewieschei 一种 , 渐新世时期。
中新海牛属Miosiren生存于中新世中期的欧洲 , 一共有Miosiren kocki 及 Miosiren canhami 两种。Miosiren kocki 全长可达3至4米 , 牠可能是螺食性 , 以獠牙于海底挖出螺类进食。
海牛亚科Trichechinae
第三纪的海牛科成员可能存活于亚马逊盆地三侧的江河一带 , 于晚中新世时期美洲中部海道的出现使得海牛亚科成员得以分布广泛 , 沿着加勒比海及热带的南美大西洋沿岸 , 直到太平洋沿岸皆有其分布 。中新世生活于海河沿岸及泻湖的Metaxytherium 出现并和海牛科产生激裂竞争因此迫使海牛科扩大生境寻找更多的新鲜水生植物 。
Sirenotherium只有Sirenotherium pirabense一种 ,可能是属于中新世早期的海牛类 , 亦可能是属于早期的儒艮类 , 这是1967年Paula Couto 根据巴西找来的两颗牙齿及颅后的节片而描述的。
Potamosiren只 Potamosiren magdalenensis一种 , 生存于中新世中期(1500万年前至1000万年前)的哥伦比亚 , 很可能是属于最早期已知的海牛亚科成员 , 跟Metaxytherium ortegense 同义。Potamosiren magdalenensis是欠缺像后来的海牛科属种具有的额外牙齿能作水平替换这特征 , 这暗示带硅酸的草本植物禾本科(Gramineae)并非其日常食物中最重要的部分。
Ribodon只有Ribodon limbatus 一种 , 生存于晚中新世至早上新世(500万年前至600万年前 ) 的阿根廷及卡罗莱纳州北部 ,可能演化出后来的海牛属Trichechus , 牙齿演化出能无限制性水平替换而这特征于现今海牛类中是存在的。
海牛科为了适应新的食物来源 , Ribodon首先拥有额外的臼齿能作水平替换 , 稍后臼齿的尺寸缩小 , 珐琅脊长度增加 , 珐琅褶变得复杂 , 排齿中牙齿数目增加 , 于海牛属Trichechus中表现出来。
海牛属Trichechus从上新世晚期或更新世(180万年前)兴起并一直延存至现在 , 海牛属Trichechus最初可能分布于多盐分的水域 , 后来发展多样化 , 于第三纪的安第斯山脉形成时扩大其生境进入亚马逊盆地并发展成亚马逊海牛 。 晚上新世时期加勒比海的儒艮类灭绝时 , 西印度海牛则迅速填补其真空地带。
安第斯山脉形成时 , 太平洋的Ribodon就这样进入亚马逊盆地并演化成现今海牛属Trichechus。亚马逊海牛Trichechus inunguis种群发展迅速 , 而且具备特有的特征 , 臼齿变得细小及复合 , 以多种水生与半水生维管束植物为食。趾甲失去 , 倍数染色体数目增加 。出生时身长体约75-85cm、10-15kg , 最大身长体重记录分别为2 . 8m及480kg , 寿命约60-70年 。
西印度海牛(Trichechus manatus ), 出现于130万年前的更新世早期 , 最初出现于佛罗里达 , 更新世晚期至全新世扩展至牙买加 , 路易斯安那州 , 俄亥俄州 , 阿肯色州 , 佛罗里达州 , 南卡罗莱纳州 , 北卡罗莱纳州 , 马里兰及新泽西州等地 。出生时身长体重分别约80至160cm及30kg , 雄性最大身长纪录分别为3 . 7m , 雄性最大身长纪录分别为4 . 1m , 最重可达1,400kg , 寿命约60~70年 。
与其它海牛相同,西印度海牛有流线形的身躯,背脊宽阔而无背鳍。头小,浑身呈灰色,但有些个体外观呈褐、红、或白色,原因可能是藻类或藤壶附着于皮肤表面。皮肤厚而紧实,表面粗糙,体毛稀疏甚至无毛。雌性体型一般较雄性略大。海牛仅具臼齿,会从上、下颚基部水平地往前移动更换。
以下是有关西印度海牛的亚种 :
1. 安地列斯海牛(Antilles Manatee, Trichechus manatus manatus)。
2. 贝克和林海牛 (Trichechus manatus bakerorum ) , 生存于晚更新世 (120,000-125,000年前) 的美国东南部 , 佛罗里达 , 北卡罗莱纳州 , 路易斯安那州等北美地域 。
3. 佛罗里达海牛(Folorida Manatee, Trichechus manatus latirostris)。
4. cf. Trichechus sp. , 一种未定的相似种 , 生存于上新世至更新世时期的巴西亚马逊盆地西部 , 跟佛罗里达海牛Trichechus manatus latirostris相比 ,安地列斯海牛Trichechus manatus manatus较类似这绝种海牛。
西非海牛(Trichechus senegalensis)于晚上新世或更新世时期由南美扩散越过大西洋至非洲 , 现分布于塞内加尔至安哥拉 , 同时西非海牛也是最少被人类研究的现存品种 。出生时身长体重分别约80至150cm及30kg雄性最大身长纪录为3 . 3m、雌性最大身长纪录为3m , 重量最大可达1,400kg , 寿命可能达60年以上。
西非海牛的外观接近其近亲西印度海牛,体型呈纺锤状,背部宽阔无背鳍,头部比例小。体色灰,皮肤厚而硬,略微粗糙,体毛稀疏。栖息于海岸的个体可能有藤壶附着。眼睛小且略微突出(与西度印度海牛不同,其眼睛较为深陷),没有外耳壳。吻部比西印度海牛肥厚而较短,可能是为了适应摄食浮水、挺水植物与生长于浅滩的植物。前肢外侧有三至四个趾甲残留。仅具臼齿,会不断地更换。
侏儒海牛Trichechus bernhardi 是近来被发现于亚马逊河流的海牛属新种 , 牠长度仅只有1.3米 及重60公斤 , 腹部有白色斑点 , 侏儒海牛Trichechus bernhardi 进食的水生植物种类也跟亚马逊海牛Trichechus inunguis 的有所不同 , 此外侏儒海牛Trichechus bernhardi也偏好较浅的急流环境 , 这和亚马逊海牛Trichechus inungui的完全相反 , 许多科学家都认为侏儒海牛Trichechus bernhardi是属于新种 , 但是于世界自然保护联盟却未被确认的。
在本文为化石网论坛精华帖,由网友秋叶月岚提供,仅供大家学习参考。
编辑:刘琮滢