
众所周知于三亿年前至少有三类主要的爬行类统治地球 , 包括下孔亚纲(Synapsida), 无孔亚纲(Anapsida)及双孔亚纲 (Diapsida) , 恐龙 , 鳄类 , 喙头蜥 , 蛇类及蜥蝪都属于双孔亚纲 (Diapsida)。巨蜥下目(Varanoidea)真正的起源时间源于晚侏罗纪 , 约一亿八千万年前。
巨蜥超科有丰富的化石记录因为牠们有高度的分化包括蛇类 , 沧龙科(Mosasauridae) , 伸龙科(Dolichosauridae) , 岸蜥科(Aigialosauridae) , 毒蜥科(Helodermatidae) , 新蜥科(Necrosauridae)及巨蜥科(Varanidae)。沧龙科(Mosasauridae) , 伸龙科(Dolichosauridae)及岸蜥科(Aigialosauridae)是海生爬行类 , 但是这些海生巨蜥超科成员于白垩纪结束时已全军覆没。
有关巨蜥类的先祖是谁至今还是不明的 , 像晚侏罗纪英国的Nuthetes destructor曾被认为是和巨蜥类有关的 , Nuthetes destructor仅只留下3.5厘米长的颌骨 , 不过后来牠被认为是属于细小的兽脚亚目恐龙。
早侏罗纪印度的印度帕卡蜥(Paikasisaurus indicus)曾被认为是巨蜥超科的先祖 , 印度帕卡蜥(Paikasisaurus indicus)只有两个颌骨节片,牙齿及髂骨的化石发现 , 印度帕卡蜥(Paikasisaurus indicus)颌骨节片非常细小 , 牙齿齿冠微曲有锋利的边缘及膨大的基部 , 牙齿基部有皱纹 , 从印度帕卡蜥(Paikasisaurus indicus)牙齿化石来看牠们看似是和巨蜥超科有关连 。不过近来的研究显示那些基部的皱纹并不是折形牙质(plicidentine) , 此外也没有其它证据显示印度帕卡蜥(Paikasisaurus indicus)是巨蜥超科的先祖 ,再说发现的化石太断续零碎也不能证明这些零碎化石是属同一个物种。
小盗蜥属(Parviraptor)有较宽及长的上腭, 较宽的翼间窝, 锋利而向后弯曲的牙齿 , 小盗蜥属(Parviraptor)额骨嗅觉部分是类似巨蜥超科 , 不过小盗蜥属(Parviraptor)牙齿没有折形牙质(plicidentine) , 牠保留了一些近祖特征显示小盗蜥属(Parviraptor)位置是和巨蜥超科先祖相当接近。小盗蜥属(Parviraptor)一共有两种包括中侏罗纪(约一亿七千万年前)英国的埃斯特斯小盗蜥(Parviraptor estesi)及美国西部的吉氏小盗蜥(Parviraptor gilmorei) , 埃斯特斯小盗蜥(Parviraptor estesi)被认为和巨蜥超科起源没关 , 至于吉氏小盗蜥(Parviraptor gilmorei)和异特龙属(Allosaurus) , 梁龙属(Diplodocus)及剑龙属(Stegosaurus)共存于当时的北美西部 , 类似巨蜥类的蜥蜴于恐龙时代已出现了。
新蜥科(Necrosauridae)
新蜥科(Necrosauridae)是一些原始的陆上巨蜥超科成员的大集会 , 活跃于晚白垩纪至始新世的北美及渐新世的欧亚大陆。牠们是已知最古老且和巨蜥类存有关系的蜥类 , 新蜥科(Necrosauridae)意思是”lizards of death” 。 Lee于1997年从分析指出新蜥科(Necrosauridae)是一类原始的类群并和沧龙类 , 毒蜥类及巨蜥类没有多大关系 , 牠们可能是并系群。
Lev Nessov于1981年描述了一块于苏联乌兹别克晚白垩纪地层发现的离散蜥类上颌骨 , Ekshmer bissektensis 就是首次被发现于苏联晚白垩纪地层的蜥类 , 鼻骨边缘陡峭高耸 , 这特征和怀俄明副萨尼瓦蜥(Parasaniwa wyomingensis)相似 , 因此Nessov认为Ekshmer bissektensis和副萨尼瓦蜥属(Parasaniwa)有关。Ekshmer bissektensis有较大的牙齿且彼此相隔较宽 , Nessov于1997年认为牠是属于新蜥科(Necrosauridae) , 但看似出现时间较晚 , Ekshmer bissektensis可能起源于科尼亚克阶(8800万年前) , Ekshmer bissektensis于新生代没有任何化石记录。
窄体副巨蜥(Paravaranus angustifrons)是生活于白垩纪蒙古 , 牠有较窄的额骨及较其它巨蜥类更大的眶部 , 并有突出的颧弓 , 这显示窄体副巨蜥(Paravaranus angustifrons)有相对较大的眼球及翼肌。Borsuk-Bialynicka 认为窄体副巨蜥(Paravaranus angustifrons)和巨蜥类起源有关 , 也有专家于1997年认为窄体副巨蜥(Paravaranus angustifrons)是巨蜥类基层成员 , 和巨蜥属(Varanus)有密切的关系。
于蒙古晚白垩纪地层也发现了另一种蜥蝪Parviderma inexacta , Parviderma inexacta只有一具不完整头骨 , 吻部及脸颊部分缺失 。Parviderma inexacta同様有着相对较大的眶部及不寻常狭窄的额骨 , Borsuk-Bialynicka于1984年认为这化石样本是属幼体的 , Parviderma inexacta头盖骨被细小的皮内成骨形成硬的外层 , 类似今天美洲中部的瘤鳞蜥属种Xenosaurus grandis。Parviderma inexacta可能是一种细小的蜥蜴 , 保存的头骨部分只有1厘米长 , 完整的话可能也不超过2厘米 , Parviderma inexacta存有七至八颗牙齿。Lee于1997年认为Parviderma inexacta是戈壁毒蜥(Gobiderma pulchrum)的姐妺群并和埃斯塔蜥属(Estesia)及毒蜥科有关。
Colpodontosaurus cracens是生活于晚白垩纪怀俄明州 , 只有两块上颌骨节片及不完整的幼长牙齿 , 两颗上颌骨节片都没有扁长的牙齿及折形牙质 , Colpodontosaurus cracens全长可能少于1米。Colpodontosaurus cracens化石缺少巨蜥类共源性(synapomorphy) , 但和原巨蜥属(Proplatynota)同样被Norell及Gao(1997年)视作属于巨蜥科。由于Colpodontosaurus cracens化石太零碎 , 更早以前Estes于1964年把Colpodontosaurus cracens视作蛇蜥下目(Diploglossa)分类不定 , 牠也可能是属于异蜥科 (Xenosauridae)或蛇蜥科(Anguidae)。Colpodontosaurus cracens牙齿有着尖吻原似巨蜥(Provaranosaurus acutus)的特征 , 有着非常狭幼的壁 。 Colpodontosaurus cracens化石形态显示牠和尖吻原似巨蜥(Provaranosaurus acutus)都是较原始的成员 , Estes(1983年)认为Colpodontosaurus cracens可能是尖吻原似巨蜥(Provaranosaurus acutus)的先祖。Colpodontosaurus cracens要是真的属于巨蜥类的话 , 牠恐怕是极原始的已知属种。
晚白垩纪怀俄明州及蒙大拿州发现的怀俄明副萨尼瓦蜥(Parasaniwa wyomingensis)属小型蜥类 , 牙齿只有2厘米长 , 仅只有颅骨节片化石发现。怀俄明副萨尼瓦蜥(Parasaniwa wyomingensis)上颌骨有较高的背突 , 有较深的吻部 , 这特征可能是独有裔征 。另一属种短钝副萨尼瓦蜥(Parasaniwa obtuse)牙齿则和怀俄明副萨尼瓦蜥 (Parasaniwa wyomingensis)有明显不同地方。
古新世蒙大纳及怀俄明州的尖吻原似巨蜥(Provaranosaurus acutus)是一种中型蜥类 , 全长少于1米。牠上颌的鼻骨边缘陡峭地上升 , 但是化石样本不完整因此难以显示上颌骨的鼻骨高度是否和怀俄明副萨尼瓦蜥(Parasaniwa wyomingensis)的相似 。尖吻原似巨蜥(Provaranosaurus acutus)有较长及苗条的牙齿 , 这可能是独有裔征的表现。尖吻原似巨蜥(Provaranosaurus acutus)牙齿有非常细小的璧 , 这显示牠具有Colpodontosaurus cracens的特征 , 尖吻原似巨蜥(Provaranosaurus acutus)下颌牙齿也同时有着怀俄明副萨尼瓦蜥(Parasaniwa wyomingensis)的特征。
始新世中期德国的凯恩始副萨尼瓦蜥(Eosaniwa koehni)头骨长达19厘米 , 全长约2米 , 有拉长的头骨及吻部 , 吻部侧旁扁压。凯恩始副萨尼瓦蜥(Eosaniwa koehni)下颌骨存有多达30颗牙齿 , 牙齿修长苗条 , 有专家认为凯恩始副萨尼瓦蜥(Eosaniwa koehni)能捕食小型哺乳类了。凯恩始副萨尼瓦蜥(Eosaniwa koehni)和其它巨蜥类关系还是不明的 , 系统发育的分析显示凯恩始副萨尼瓦蜥(Eosaniwa koehni)是较原始的巨蜥类 , 和沧龙类 , 库尼亚蜥属(Coniasaurus)及副巨蜥属(Paravaranus)属同一进化支中。
新蜥属(Necrosaurus)生存的时代为古新世至始新世 , 头骨能长达7厘米。新蜥属(Necrosaurus)有广大的颌骨内收肌并延至头盖 , 和颅顶骨顶部相连形成矢状嵴(sagittal crest)。新蜥属(Necrosaurus)有两种包括法国的Necrosaurus cayluxi及德国的Necrosaurus eucarinatus , Necrosaurus eucarinatus有较多牙齿, 牙齿数目能多达18颗 , Necrosaurus cayluxi仅只有13颗牙齿。Necrosaurus eucarinatus颅顶骨没有矢状嵴(sagittal crest) , 这可能只是显示年龄阶段不同罢了。
毒蜥科
毒蜥科要追溯至晚白垩纪, 大约8000万年前 , 牠们曾生活于欧洲, 亚洲及北美 , 但于始新世以后仅只在北美取得分布。至于毒蜥属这世系的历史则长达二千万年。
最古老的巨蜥超科成员是来自美国犹他州白垩纪地层 , 只有一具不完整的右上颌化石 , 这具化石现被认为是妮索侏毒蜥(Primaderma nessovi), 妮索侏毒蜥(Primaderma nessovi)生存于阿尔布阶(Albian)至森诺曼阶(Cenomanian)。侏毒蜥属(Primaderma)上颌骨具有前上颌骨突并于上颌牙齿前缘及后缘形成背突边及幼条纹 ,侏毒蜥属(Primaderma)是最古老的巨蜥超科成员 , 从分类学上来看牠们是类似毒蜥科的巨蜥超科成员 , 侏毒蜥属(Primaderma)牙齿有着巨蜥超科成员数个祖征但没有形成毒液沟。从妮索侏毒蜥(Primaderma nessovi)的头骨来看侏毒蜥属(Primaderma)是具高度掠食性并有足够能力捕杀较大的猎物。妮索侏毒蜥(Primaderma nessovi)的发现显示巨蜥科及毒蜥科的起源大概发生在一亿年前。
戈壁毒蜥(Gobiderma pulchrum)是生活于晚白垩纪蒙古的中型蜥类 , 头骨长约5厘米 , 牠仅只有头骨及下颌化石发现 , 全长推测可能少于1米及达1米左右。戈壁毒蜥(Gobiderma pulchrum)头骨后边较宽 , 不过牠的嗅觉系统和以后的巨蜥类有所不同。戈壁毒蜥(Gobiderma pulchrum)的头盖及面颊保留了皮内成骨(Osteoderms), 这特征类似毒蜥科 , 而且两者的皮内成骨结构也有相似。戈壁毒蜥(Gobiderma pulchrum)的下颌和毒蜥科相似但前者有较小的喙突 , 戈壁毒蜥(Gobiderma pulchrum)有较锐利的牙齿 , 牙齿间有宽阔的间隔 , 下颌比上颌略长。对于戈壁毒蜥(Gobiderma pulchrum)和其它巨蜥类的关系 ,1997年 Lee认为戈壁毒蜥(Gobiderma pulchrum)和副巨蜥属(Paravaranus)有密切的关系 , 但后来受到质疑 , 2000年 Alifanov也不认同戈壁毒蜥(Gobiderma pulchrum)和副巨蜥属(Paravaranus)有关连 , 他认为戈壁毒蜥(Gobiderma pulchrum)应是和新蜥属(Necrosaurus)关系较近 , 但却没有公布原因何在。
博格副毒蜥(Paraderma bogerti)只有颅骨节片化石发现 , 牠是生活于晩白垩纪马斯特里赫特阶的怀俄明州。Estes(1983)并不确定博格副毒蜥(Paraderma bogerti)和毒蜥科的关系 , 但是1997年Norell及Gao通过对化石的分析已确定牠是属毒蜥科。博格副毒蜥(Paraderma bogerti)有短而宽的吻部 , 具多边型 , 有孔的皮内成骨和上颌融合。博格副毒蜥(Paraderma bogerti)牙齿内也存有毒沟显示毒蜥科于晚白垩纪时期已在美国西部取得分布。从博格副毒蜥(Paraderma bogerti)颅骨节片化石推算博格副毒蜥(Paraderma bogerti)全长相当于现存的毒蜥属及者要略大一些 。
于新生代毒蜥科取得空前的发展 , 加利欧毒蜥(Eurheloderma gallicum)的发现显示毒蜥科并不局限于北美及亚洲 , 加利欧毒蜥(Eurheloderma gallicum)化石被发现于法国始新世岩层上 , 牠的头骨长达7至8厘米 , 较现存的美国毒蜥(Heloderma suspectum)略大 , 颅顶骨也较毒蜥属的要长。不过加利欧毒蜥(Eurheloderma gallicum)牙齿的毒腺沟没现存的毒蜥属那么发达及完善 , 较大的颌骨内收肌显示加利欧毒蜥(Eurheloderma gallicum)更依赖咬力捕杀猎物 , 相反并不依赖毒液。加利欧毒蜥(Eurheloderma gallicum)有呈多边形纹饰的皮内成骨 , 和毒蜥属相似。于怀俄明州晚古新世地层也发现类似加利欧毒蜥(Eurheloderma gallicum)的颅顶骨化石 , 这化石可能是属于亚成体的。
洛氏毒蜥(Lowesaurus matthewi)生存于渐新世中期至中新世早期的内布拉斯加及科罗拉多州, 牠是中小型蜥蝪 , 洛氏毒蜥(Lowesaurus matthewi)头颅的皮内成骨相对性要较毒蜥属的来得长。洛氏毒蜥(Lowesaurus matthewi)额骨呈三角形 , 形态显示洛氏毒蜥(Lowesaurus matthewi)要较毒蜥属原始 , 不过洛氏毒蜥(Lowesaurus matthewi)牙齿前边已有明显的毒腺沟。
毒蜥属(Heloderma)化石曾被发现于内华达州 , 新墨西哥州及德克萨斯州 , 于内华达州发现的化石属于美国毒蜥(Heloderma suspectum), 时期是一万年至八千年。于德克萨斯州发现的德克萨斯毒蜥(Heloderma texana)是属早中新世 , 头骨要较现存的毒蜥属种来得平坦 , 德克萨斯毒蜥(Heloderma texana)的体型也较现存的属种细小 , Pregill et al.于1986年总结德克萨斯毒蜥(Heloderma texana)是墨西哥毒蜥(Heloderma horridum)及美国毒蜥(Heloderma suspectum)的姐妹群。Estes于1983年指出佛罗里达中新世地层发现的毒蜥属未定种(Heloderma sp?)显示毒蜥科活动局限于雁列山至大西洋海岸。
巨蜥科和毒蜥科兴起的时间大概差不多 , 像蒙古的蒙古似萨尼瓦蜥(Saniwides mongoliensis)就是生活于8000万年前 , 蒙古似萨尼瓦蜥(Saniwides mongoliensis)外型上和现存的巨蜥属很相似 , 有非常低 , 平坦及相对宽广的吻部 , 并有着鸭嘴的外型 。蒙古似萨尼瓦蜥(Saniwides mongoliensis)头骨长度少于5厘米显示蒙古似萨尼瓦蜥(Saniwides mongoliensis)全长小于1米 。蒙古似萨尼瓦蜥(Saniwides mongoliensis)颌骨内收肌和颅顶骨侧面相连 , 蒙古似萨尼瓦蜥(Saniwides mongoliensis)有略高的颅顶骨 , 有较大的内收肌显示蒙古似萨尼瓦蜥(Saniwides mongoliensis)有较大的咬力。从头骨形态来看蒙古似萨尼瓦蜥(Saniwides mongoliensis)较毒蜥科存有更多祖征。
长吻原巨蜥(Proplatynota longirostrata)是一种中型大小的巨蜥类 , 头骨只有4厘米长 , 长吻原巨蜥(Proplatynota longirostrata)的头骨修长而苗条 , 和戈壁毒蜥(Gobiderma pulchrum)及副巨蜥(Paravaranus angustifrons)同样没有侧向突出的颧弓。
东方艾俄洛斯蜥(Aiolosaurus oriens)是近来才被描述的白垩纪巨蜥科成员 , 牠的化石样本不完整 , 头骨只有4至5厘米 , 全长应少于1米。东方艾俄洛斯蜥(Aiolosaurus oriens)有相对较深的上颌孔 , 有多达9至10个锋利而后弯的牙齿及少量折形牙质(plicidentine)。东方艾俄洛斯蜥(Aiolosaurus oriens)和Cherminotus longifrons同样有一个泪腺孔 , 和蒂尔玛蜥属(Telmasaurus), 萨尼瓦蜥属(Saniwa) , 拟毒蜥属(Lanthanotus)及巨蜥属(Varanus)则具两个泪腺孔 , 这显示东方艾俄洛斯蜥(Aiolosaurus oriens)和Cherminotus longifrons要比以上四个属来得原始。
于蒙古南戈壁(Omnogo’)晚白垩纪Djadoktha组发现了巨蜥科新属种骨尔敖包蜥(Ovoo gurval), 骨尔敖包蜥(Ovoo gurval)和在蒙古的乌哈托喀(Ukhaa Tolgod)发现的细小巨蜥科属种东方艾俄洛斯蜥(Aiolosaurus oriens)相似 , 骨尔敖包蜥(Ovoo gurval)有着许多东方艾俄洛斯蜥(Aiolosaurus oriens)的特征 , 此外骨尔敖包蜥(Ovoo gurval) 头骨顶上有一对不寻常的新生变体的骨化成骨并覆盖额鼻区边缘。此外骨尔敖包蜥(Ovoo gurval)这个新生变体的骨化特征和其它已知的有鳞目成员大不相同 , 此外皮内成骨(Osteoderms)也和一些巨蜥类有明显分别。
晚白垩纪蒙古的Cherminotus longifrons是一种体型细小的蜥类 , 头骨少于3厘米 , Cherminotus longifrons上颌有五颗彼此间隔宽阔的圆锥形牙齿 。古生物学家最初认为Cherminotus longifrons是拟毒蜥(Lanthanotus borneensis)的姐妹群 , 但是后来古生物学家对这结论产生了疑问 , 他们认为Cherminotus longifrons看似是和拟毒蜥(Lanthanotus borneensis)没有任何关系 。
根据巨蜥科及牠们的近亲毒蜥属 (Heloderma)及拟毒蜥科(Lanthanotidae)至少在九千万年前源于亚洲北部 , 已知最古老的巨蜥科属种是来自蒙古包括谷氏蒂尔玛蜥(Telmasaurus grangeri), 蒙古似萨尼瓦蜥(Saniwides mongoliensis)及蒙古埃斯塔蜥(Estesia mongoliensis) , 牠们的外型和现存的巨蜥属种有些相似。
蒙古埃斯塔蜥(Estesia mongoliensis)生活于晚白垩纪的蒙古 , 从牠的头骨侧面来看蒙古埃斯塔蜥(Estesia mongoliensis)头骨和今天的巨蜥属种很相似 , 但有较低及钝而宽的吻部。蒙古埃斯塔蜥(Estesia mongoliensis)牙齿具有沟槽显示蒙古埃斯塔蜥(Estesia mongoliensis)可能会像吉拉毒蜥(Heloderma suspectum)般把毒液注入猎物体内 , 不过现在对这些史前蜥类和毒蜥属 (Heloderma)及巨蜥科关系如何还是不明的。蒙古埃斯塔蜥(Estesia mongoliensis)看似和今天的巨蜥属种非常相似 , 但是Norell及Gao把蒙古埃斯塔蜥(Estesia mongoliensis)视作毒蜥类成员 , 他们认为蒙古埃斯塔蜥(Estesia mongoliensis)和毒蜥属(Heloderma)关系要较接近得多 , Lee于1997年也把蒙古埃斯塔蜥(Estesia mongoliensis)划作和毒蜥科较接近。
谷氏蒂尔玛蜥(Telmasaurus grangeri)的化石相当不完整 , 吻部也缺失 , 牠的头骨大概长达7至8厘米 。研究分析显示谷氏蒂尔玛蜥(Telmasaurus grangeri)和萨尼瓦蜥属(Saniwa) , 拟毒蜥属(Lanthanotus)及巨蜥属(Varanus)有密切的关系。
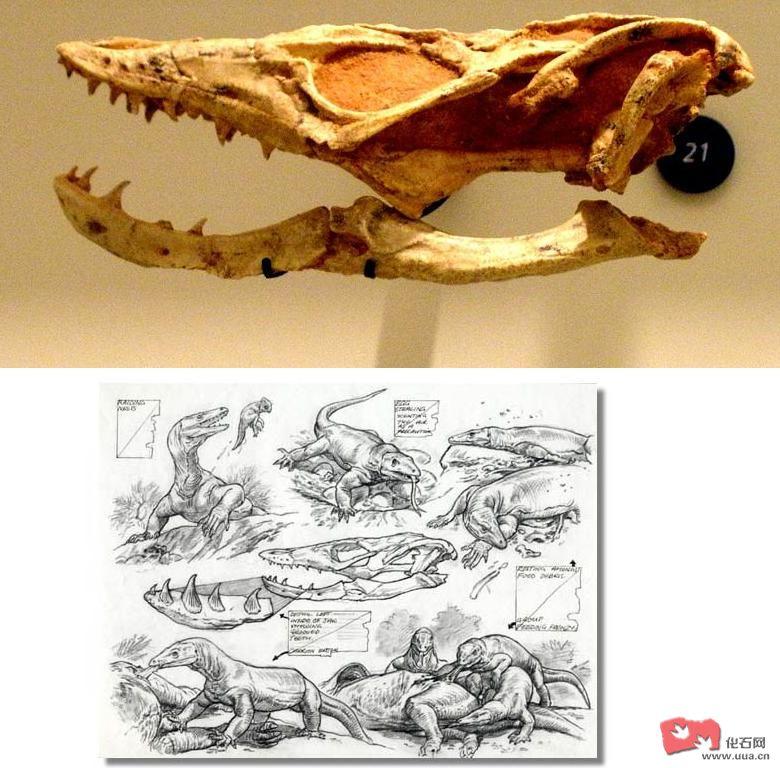
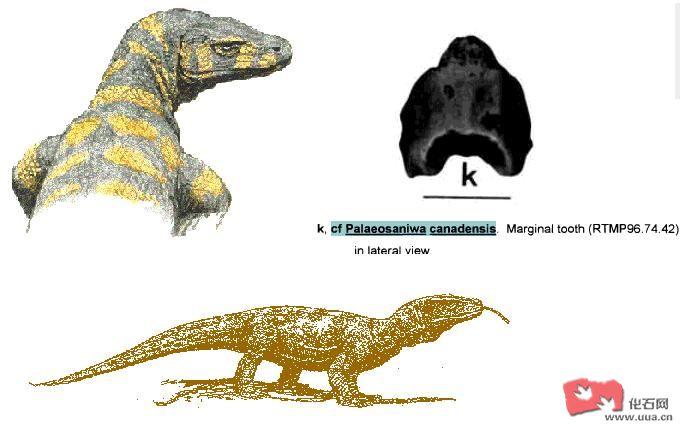
加拿大古萨尼瓦蜥(Palaeosaniwa canadensis)是一种暂定作巨蜥科的史前蜥类 , 牠的化石被发现于加拿大艾伯塔省(Alberta)及美国怀俄明州 , 大多学者认同北美的巨蜥科属种是源于亚洲的 。加拿大古萨尼瓦蜥(Palaeosaniwa canadensis)是生存于七千万年前的北美 , 全长可达2.4米。[一说加拿大古萨尼瓦蜥(Palaeosaniwa canadensis)最大可达10-12呎及重达150公斤]加拿大古萨尼瓦蜥(Palaeosaniwa canadensis)有长而后弯的牙齿 , 牙齿长有锯缘并长有类似蒙古埃斯塔蜥(Estesia mongoliensis)的牙沟槽 , 尽管加拿大古萨尼瓦蜥(Palaeosaniwa canadensis)长得已和现在的巨蜥非常相似 , 但划进巨蜥科还是存有疑问的。
已灭绝的萨尼瓦蜥属(Saniwa)和巨蜥属(Varanus)关系较密切 , 萨尼瓦蜥属(Saniwa)是生活于始新世至渐新世北美及欧洲的巨蜥科成员 , 全长能达1米 , 这个属有数个已命名的种 。萨尼瓦蜥属(Saniwa)像大多数陆上巨蜥类成员一样有着相对较大的头部。萨尼瓦蜥属(Saniwa)形态上和现存的巨蜥相似 ,牠们保留了许多巨蜥属的祖征包括细小的前上颌骨及六颗前上颌骨牙齿 , 尾近侧有前后膨大的神经棘。
始新世北美的肯塞萨尼瓦蜥(Saniwa ensidens)有较长及狭窄的头骨 , 并具有下颌内关节(intramandibular joint)。 肯塞萨尼瓦蜥(Saniwa ensidens)有多达6颗前上颌骨牙齿 , 24颗上颌骨牙齿 , 有至少22颗牙齿每颗牙齿都异常锋利尖锐并向后弯曲 , 基部膨胀并具有脊纹。至于始新世怀俄明州的Saniwa agilis可能是肯塞萨尼瓦蜥(Saniwa ensidens)的同物异名 , Saniwa agilis脊椎长度只有10mm。始新世怀俄明州的粗厚萨尼瓦蜥(Saniwa crassa)有较厚重的脊椎结构显示粗厚萨尼瓦蜥(Saniwa crassa)有较大的体型 , 体型大小相当于草原巨蜥(Varanus exanthematicus),但是脊椎中枢的髁前结构和其它萨尼瓦蜥属种没有多大明显不同。于当时怀俄明州也存在更大的属种---格兰迪萨尼瓦蜥 (Saniwa grandis) , 但是格兰迪萨尼瓦蜥(Saniwa grandis)这个种名可能不是有效的 , 牠可能只是一个特大的个体罢了 , 格兰迪萨尼瓦蜥(Saniwa grandis)保留了厚重的脊椎。至于怀俄明州及犹他州始新世地层发现的Saniwa paucidens这个种名也常不被认为是有效学名的。粗厚萨尼瓦蜥(Saniwa crassa)也只是肯塞萨尼瓦蜥(Saniwa ensidens)的同物异名。从分析上显示肯塞萨尼瓦蜥(Saniwa ensidens)是巨蜥属(Varanus)的姐妹群。
始近世南加利褔亚的布氏萨尼瓦蜥(Saniwa brooksi)和肯塞萨尼瓦蜥(Saniwa ensidens)两者分布上没有重叠 , 两者骨骼也明显不同。美国北达克州晚始新世Chadron组发现的最后萨尼瓦蜥(Saniwa edura)体型可能相当或比肯塞萨尼瓦蜥 (Saniwa ensidens)略大 , 最后萨尼瓦蜥(Saniwa edura)牙齿也较肯塞萨尼瓦蜥 (Saniwa ensidens)的略大。
菲氏萨尼瓦蜥(Saniwa feisti)生活于始新世的西欧 , 化石被发现于德国 , 体长约60厘米 , 有较短的颈部 , 颚骨有多达20颗牙齿 , 牙齿数目和肯塞萨尼瓦蜥 (Saniwa ensidens)的不相同。从分析研究显示菲氏萨尼瓦蜥(Saniwa feisti)是巨蜥科的基层成员。
至于比利时发现的奥尔沙萨尼瓦蜥(Saniwa orsmaelensis)躯干椎骨已和菲氏萨尼瓦蜥(Saniwa feisti)没明显分别。奥尔沙萨尼瓦蜥(Saniwa orsmaelensis)生存于晚古新世 , 牠是最古老的萨尼瓦蜥属种。
于蒙古晚白垩纪至始新世地层曾有发现过萨尼瓦蜥属未定种(Saniwa sp)的化石 , 但这些都只是脊椎化石发现。
于内布拉斯加及怀俄明州渐新世中期地层也发现了一些欵似萨尼瓦蜥属(Saniwa)的椎骨化石 , 这些化石并不能完全肯定属于萨尼瓦蜥属(Saniwa) , 化石距今3200万年前 , 以后再没有任何疑似萨尼瓦蜥属(Saniwa)的化石记录了。不过巨蜥类却于旧大陆繁荣过来 , 而毒蜥类则于新大陆发展 , 牠们分别盘据东西半球。
巨蜥属(Varanus)于蒙古始新世地层已有化石记录 , 但是化石太零碎断续因此难以定种的。于中新世时期大幅扩散 , 欧洲 , 哈萨克 , 东非都有巨蜥属的化石记录。于2000万年前有一种露辛加巨蜥(Varanus rusingensis)栖息于露辛加岛(Rusinga Island )及肯尼亚其它地区 ,露辛加巨蜥(Varanus rusingensis)形态和现存的非洲巨蜥属种尼罗河巨蜥(Varanus niloticus), 草原巨蜥(Varanus exanthematicus)及白喉巨蜥(Varanus albigularis)非常相似 。 露辛加巨蜥(Varanus rusingensis)可能主要以软体动物为食 , 生活于森林并具有类似尼罗河巨蜥(Varanus niloticus)的半水生习性 , 露辛加巨蜥(Varanus rusingensis)全长至少达2米。
霍夫曼巨蜥(Varanus hofmanni )是已知最古老的欧洲巨蜥属种 , 生存于距今一千三百万年前的西欧 , 化石被发现于法国 , 西班牙及德国 。 另一种被发现于摩尔达维亚中新世海床的Varanus tyrasiensis椎骨长度达170mm , 有专家认为Varanus tyrasiensis和霍夫曼巨蜥(Varanus hofmanni)是同物异名。和霍夫曼巨蜥(Varanus hofmanni)同时期出现并有密切关系的还有西班牙及葡萄牙的克托尼俄斯伊比利亚巨蜥(Iberovaranus catalonicus)及哈萨克斯坦的普罗宁巨蜥(Varanus pronini)。克托尼俄斯伊比利亚巨蜥(Iberovaranus catalonicus)仅只有椎骨化石发现 , 椎骨和毒蜥科的相似 , 于葡萄牙也发现了牠的相似种化石但和正模标本有些不同 , 可能只是个体变异罢了。普罗宁巨蜥(Varanus pronini)化石被发现于中新世中期地层 , 距今1300万年前 , 椎体中枢有较宽的髋臼且髋臼占去中枢长度的74% , 这特征和其它巨蜥属种很不相同。
马拉松巨蜥(Varanus marathonensis)出现时间是500万年前 , 在希腊 , 匈牙利及土耳其都有化石记录 , 据说于意大利更新世地层也有马拉松巨蜥(Varanus marathonensis)的化石记录 , 但这记录还没未被确认 。马拉松巨蜥(Varanus marathonensis)的中枢要较霍夫曼巨蜥(Varanus hofmanni)来得长 , Orlov 及 Tuniev 认为马拉松巨蜥(Varanus marathonensis)和现存的荒漠巨蜥(Varanus griseus)及生活于500万年前塔吉克斯坦的Varanus darevskii有非常密切的关系。Varanus darevskii体型较荒漠巨蜥(Varanus griseus)来得细小 , 但是Varanus darevskii有较高的头骨 , 较短的吻部 , 较大及锋利的牙齿。甚至曾有专家于1986年假定Varanus darevskii可能是荒漠巨蜥(Varanus griseus)的先祖 。古生物学家把马拉松巨蜥(Varanus marathonensis) , Varanus darevskii和现存的荒漠巨蜥(Varanus griseus)三者划作Psammosaurus这个亚属内 , 而荒漠巨蜥(Varanus griseus)至目前还没有任何化石记录。
西蒙巨蜥(Varanus semjonovi )是被发现于乌克兰的中新世属种 , 全长约50至70厘米 ; 伦古巨蜥(Varanus lungui)是生活于中新世的摩尔达维亚 , 全长约1米 , 伦古巨蜥(Varanus lungui)和印度巨蜥(Varanus indicus)关系要较密切但牠要较印度巨蜥(Varanus indicus)原始得多。同时期还有印度的西哇巨蜥(Varanus silvalensis) , 西哇巨蜥(Varanus silvalensis)全长约3米。Lydekker根据水巨蜥(Varanus salvator)的骨骼比例来判断西哇巨蜥(Varanus silvalensis)长约3.4米至3.7米。
已灭绝的郝益巨蜥(Varanus hooijeri)生存于上新世时期(500万年前以后)的危地马拉 , 郝益巨蜥(Varanus hooijeri)头骨有结实及钝的牙齿 , Estes认为郝益巨蜥(Varanus hooijeri)和现存的葛氏巨蜥 (Varanus olivaceus)或尼罗河巨蜥(Varanus nitoticus)关系要较密切 , 郝益巨蜥(Varanus hooijeri)全长相当于尼罗河巨蜥(Varanus nitoticus)。郝益巨蜥(Varanus hooijer)有膨胀及不锋利的臼型齿 , 这可能是独有衍征(autapomorphy)。
现在对巨蜥科于澳洲的历史还是一无所知的 , 已知澳洲最古老的巨蜥科化石来自1千万年前 , 于新南威尔士州便发现了类似眼斑巨蜥(Varanus giganteus)的椎骨化石 , 化石所属时期少于200万年前 。Baverstock et al于1994年指出巨蜥科于2000万年以后才从亚洲东南部进入澳洲 , 世界各地都存有中型至大型的巨蜥科属种 , 但仅只有少量属种全长才能达3米 , 巨型及侏儒的巨蜥科属种都在澳洲共同进化。于南澳洲发现了一些未被描述的早期巨蜥科属种 , 这些化石都属于500万年前 。于昆士兰发现的伊塔巨蜥(Varanus emeritus), 只有肱骨及胫骨化石发现 , 伊塔巨蜥(Varanus emeritus)化石和黄点巨蜥(Varanus salvadorii)的相似 , 从伊塔巨蜥(Varanus emeritus)的化石推测全长达3米或4米。
恐巨蜥(Varanus dirus)被发现于昆士兰Darling Downs的更新世地层 , 仅只有离散的牙齿化石发现 , 恐巨蜥(Varanus dirus)时常被认为是古巨蜥(Megalania prisca)的同物异名 。于昆士兰东南部Chinchilla的早上新世沉积层也发现了恐巨蜥(Varanus dirus)的颚骨, 三颗牙齿化石及长达2.1米的脊椎化石 , 从颚骨形态来看恐巨蜥(Varanus dirus)和黄点巨蜥(Varanus salvadorii)的非常相似 , 但是恐巨蜥(Varanus dirus)体型要较黄点巨蜥(Varanus salvadorii)大两倍 , 黄点巨蜥(Varanus salvadorii)体长最大能达244 cm。近来对恐巨蜥(Varanus dirus)的研究还是存有争论 , 有专家认为恐巨蜥(Varanus dirus)是黄点巨蜥(Varanus salvadorii)的特大个体 , 有的专家则认为只是古巨蜥(Megalania prisca)的幼体 , 也有专家认为是独立种 , 由于缺乏个体骨骼发育上的数据因此无法对此作太多深入了解。
晚更新世的古巨蜥[Megalania prisca ( Varanus prisca) ]是目前已知陆上最巨型的蜥蝪 , 成年古巨蜥(Megalania prisca)全长最大可达7米或以上及重量超过600kg , 古巨蜥(Megalania prisca)于澳洲大幅扩散 , 于新南威尔士州(New South Wales) , 昆士兰及南澳洲也有古巨蜥(Megalania prisca)的残骸发现 , 古巨蜥(Megalania prisca)一直延存至2万5千年便完全灭绝, 古巨蜥(Megalania prisca)被认为以袋鼠及袋熊等有袋类为食 , 古巨蜥(Megalania prisca)也被认为会袭击早期移居澳洲的人类。
现存巨蜥属有多达46种分布于非洲 , 亚洲及澳大拉西亚 , 有专家认为现存属种的共同先祖于4500万年前出现 , 在现存属种中尼罗河巨蜥 (Varanus niloticus)于乍得的晚中新世至上新世交界组份已有化石记录。于印度更新世地层也发现了现存的孟加拉巨蜥(Varanus bengalensis)化石 。一些现存的巨蜥属种于更新世时期的澳洲已存在了 , 在新威尔士发现了疑似眼斑巨蜥(Varanus giganteus)的化石 , 于澳洲南部维多利亚的洞穴也发现了砂巨蜥(Varanus gouldii)的化石 , 于澳洲南部也同时发现了饰纹巨蜥(Varanus varius)的化石 , 眼斑巨蜥(Varanus giganteus)体长最大能达1.9米 , 饰纹巨蜥(Varanus varius)体长最大可达2米。这些化石记录显示巨蜥属种在晚冰河时期分布于南部及东部气候较温和的地区 , 时期是14000年前至3000年前。
现在科莫多巨蜥 (Varanus komodoensis)仅分布于爪哇至澳洲之间的东印度尼西亚 , 过去一直认为科莫多巨蜥 (Varanus komodoensis)是从400万年前从来自澳洲的先祖分化过来 , 不过于澳洲昆士兰发现了科莫多巨蜥 (Varanus komodoensis)的化石, 化石属上新世时期 ,这些化石显示科莫多巨蜥 (Varanus komodoensis)是从澳洲演化过来并向印度尼西亚扩散 , 于澳洲大陆发现的科莫多巨蜥 (Varanus komodoensis)化石显示牠们起源于380万年前的澳洲大陆。
于印度尼西亚弗洛勒斯岛(Flores)也发现了早更新世时期科莫多巨蜥 (Varanus komodoensis)的化石 , 过去90万年间科莫多巨蜥 (Varanus komodoensis)体型在印度尼西亚弗洛勒斯岛一直鲜有改变 。于爪哇岛同样发现了更新世中期的科莫多巨蜥相似种(Varanus sp. cf. V. komodoensis)和水巨蜥(Varanus salvator)的化石 , 该科莫多巨蜥相似种(Varanus sp. cf. V. komodoensis)体型及形态上和科莫多巨蜥 (Varanus komodoensis)接近 , 大概能总结科莫多巨蜥 (Varanus komodoensis)是从早上新世澳洲演化 , 接着向西扩散到达印度尼西亚弗洛勒斯岛(Flores) , 最后进入爪哇。
于澳洲中部发现的更新世巨蜥属化石 , 体型上介乎于科莫多巨蜥 (Varanus komodoensis)及古巨蜥(Varanus prisca)之间 , 可能和帝汶岛发现的巨蜥属未定种(Varanus sp)同种 , 至于帝汶岛及澳洲岛之间的巨蜥种群只能待更多化石记录才能了解。
于爪哇早更新世地层也发现了水巨蜥(Varanus salvator) 的背椎 , 骶椎及尾椎 , 从这些椎骨来看牠们有着现存的水巨蜥(Varanus salvator)个体的变异 , 其中有三个背椎维持于科莫多巨蜥(Varanus komodoensis )的下限值 , 最初这些化石样本被认为是属于科莫多巨蜥(Varanus komodoensis ) , 但后来被认为是属于非常巨大的水巨蜥(Varanus salvator)个体 , 这些骨骼化石和水巨蜥(Varanus salvator)的都很相似 , 但要较科莫多巨蜥(Varanus komodoensis )及西哇巨蜥(Varanus sivalensis) 细长 。 这些早更新世的水巨蜥(Varanus salvator)个体比下图的现存水巨蜥个体大出20%以上 , 而下图的现存水巨蜥个体比现在最大的水巨蜥(Varanus salvator)个体要小15–20% , 现存水巨蜥最大记录是3.21米。
Varanus bolkayi 是生存于200万年前的爪哇及帝汶岛 , 牠可能是现存水巨蜥 (Varanus salvator)的同物异名。Varanus bolkayi仅只有两个长而苗条的椎骨 , 最大的长达2.7厘米 , 显示牠是大型蜥类 , 但不算是非常大的巨蜥属种。
西哇巨蜥(Varanus sivalensis)背椎化石样本(NHMR739)则要略大于上述的现存水巨蜥个体 , 但是化石样本(NHMR739)大小维持于爪哇早更新世地层发现的水巨蜥(Varanus salvator)个体水平 , 西哇巨蜥(Varanus sivalensis)背椎化石样本(NHMR739)大概相当于科莫多巨蜥(Varanus komodoensis )的下限值。
(在本文为化石网论坛精华帖,由网友两魏争雄提供,仅供大家学习参考。编辑:刘琮滢)