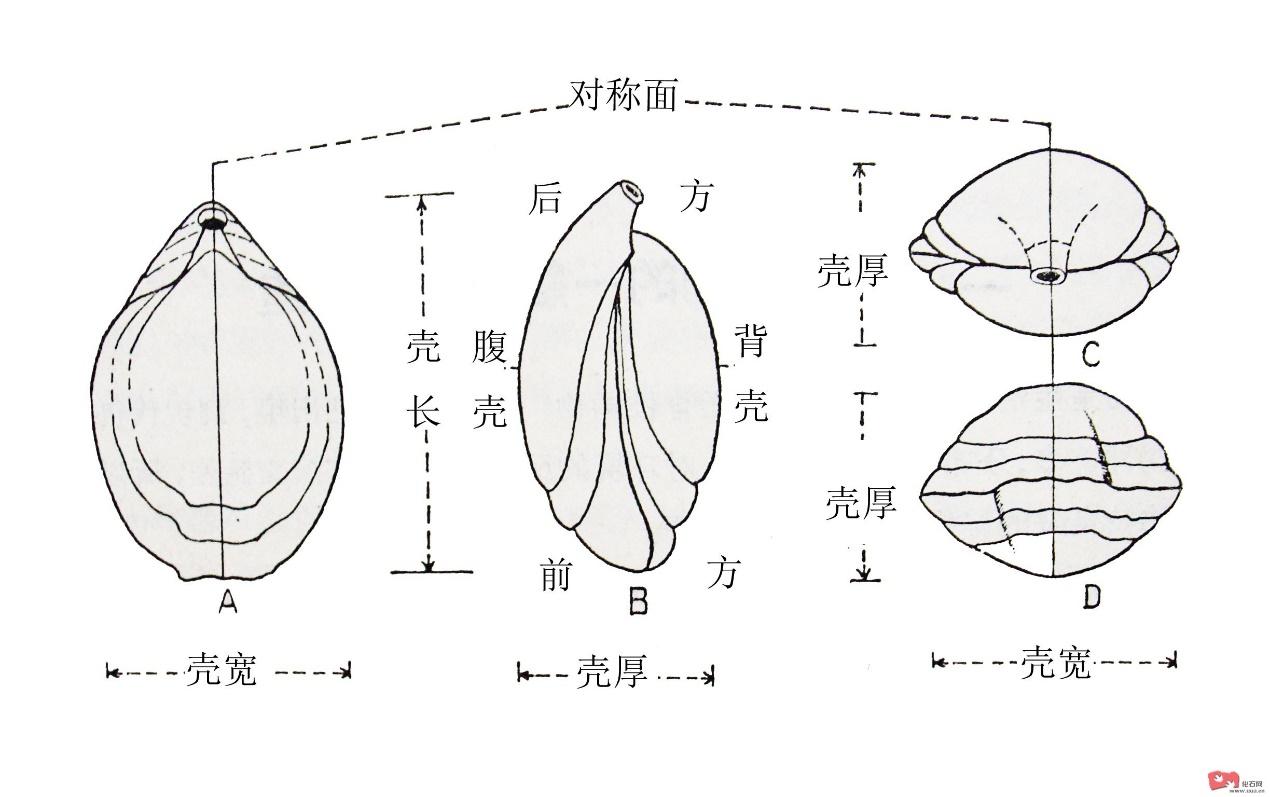
太原太原组王冠上的宝石——Schwagerina蜓属与Quasifusulina蜓属
很久以前不记得是在哪里看到的文字,大意说石炭纪后期的浅海出现了一种史无前例的生物——蜓 这种单细胞生物演化迅速,其对古地理的指示作用因为其比腕足还要敏感的环境应激而显得独一无二,从石炭纪开始,很久以前不记得是在哪里看到的文字,大意说石炭纪后期的浅海出现了一种史无前例的生物——蜓.......
这种单细胞生物演化迅速,其对古地理的指示作用因为其比腕足还要敏感的环境应激而显得独一无二,从石炭纪开始,到二叠纪末期,突然灭绝。
这个字怎么写呢?字库里没有这个字,以至于所有文献(除了专门印刷的)到这个字就打个方框表示写不出来,要么就是写作“竹蜓”.....
真是种奇怪的生物啊
明明只有一个细胞,却学软体动物整个壳;
明明只有一个细胞,却像菊石一样有多个房室;
明明只有一个细胞,却做了石炭纪标准化石的头把交椅;
明明只有一个细胞,却消失在2亿年前隆起的海床;
明明只有一个细胞.......
我相信这个东西是大家陌生的,就像这微体版块一样,只能引起大家好奇而迷惑的目光。
这个字正确的写法及读音是这样的
蜓:
有孔虫纲
蜓目 Order Fusulinda
蜓超科 Super-Family Fusulinda
此次挖掘地位于太原东山,与作为标准地层的西山相聚十公里左右,与西山一样也是太原市区电煤供应区,此地地层较之西山同层位都薄不少,是时间精力都有限的FF挖掘海路交互相沉寂海相化石的理想场所。
1983年的张志存依据蜓化石种属分异度把西山地层分为Tritictes simplex(C2)带和Pseudoschwagerina(C3)带,其中Pseudoschwagerina带被分为四个亚带,东山地区由于地层相应较薄,这四个带的分异就不是那么明显,石灰岩的层数也不如西山多。Schwagerina蜓在西山四带中都有分布,但从同层位腕足化石的种属来看,可以初步估计出该层层应为太原组上部,石炭-二叠纪分界面处,相应于1983年的划分,此次获得的Schwagerina和Qusifusulina的两个两个种应该属于太原组上部第二个亚带:Schwagerina cervicalis带。
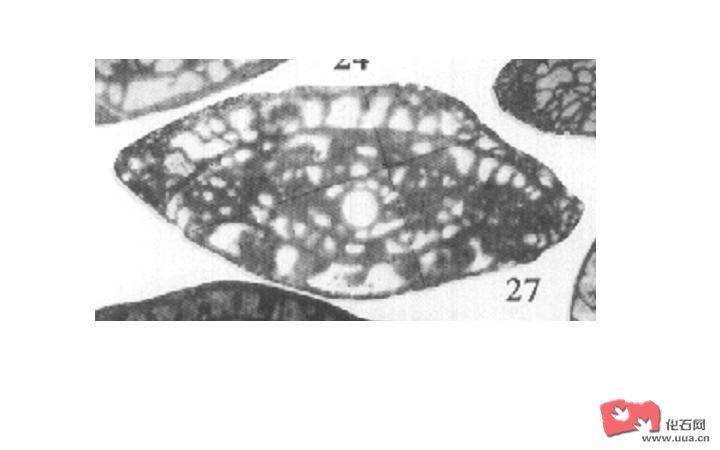
石炭纪-二叠纪的蜓通常具有很厚的蜂巢层和强烈的隔壁褶皱,如下图是Schwagerina 属elkoensis H&T种:
环绕成圈的层就是隔壁,也称旋壁,可见很黑很厚,因为它是又致密层和蜂巢层叠合而成的,中间的孔称作初房,是有孔虫的第一个房室,两边乱麻麻的黑线是隔壁褶皱。
下面就是在东山挖到的Schwagerina属的显微照片:
上图为zhangtensor用放大镜所拍,一格一毫米,偏心旋切面
此为旋切面
此为偏心旋切面20倍照片,旋壁明显厚于轴向的隔壁,并且隐约可见褶皱,此标本被粘过
此为一端的隔壁褶皱
同上
以下是五十倍下的较小个体,同为Schwagerina属
旋切
旋切
这也是个较大个体,个人认为是Schwagerina属旋切面,但是资料显示存在Sphaeroschwagerina(伪希瓦格蜓属)是球形,但同时华北型地层普遍缺失改属(频繁海退引起),这个问题请还请foraminifera版主赐教。
接下来是另一属:
Quasifusulina
改属与Schwagerina混生,但数量上居多,个体细长,有8mm左右,实体两端膨大,应该是轴积形成:
以下为实体二十倍显微:
突起部位为轴积痕迹
Quasifusulina属与Schwagerina属有着鲜明的差异,首先它的个体纤长,其隔壁厚度不及Schwagerina属,但隔壁褶皱比Schwagerina更甚,尤其在两端,并且还有及其明显的轴向次生堆积---轴积
上图是Qusifusulina 属 gracilis sheng种
上两图为偏心旋切面,其中细密的孔就是两端强烈的隔壁褶皱,中间实心部分为轴积,
上图20倍,下图为50倍,可见隔壁
下面这两张是偏心斜旋切面:
同样可见轴积与隔壁褶皱
(本文为化石网论坛精华帖,由网友johnnashzhang提供,仅供大家学习参考。)
编辑:常君滢
2022-11-24