狮虎的咬力,咬肌,肌肉和眼睛大小和剑齿形掠食者的下颌骨强度
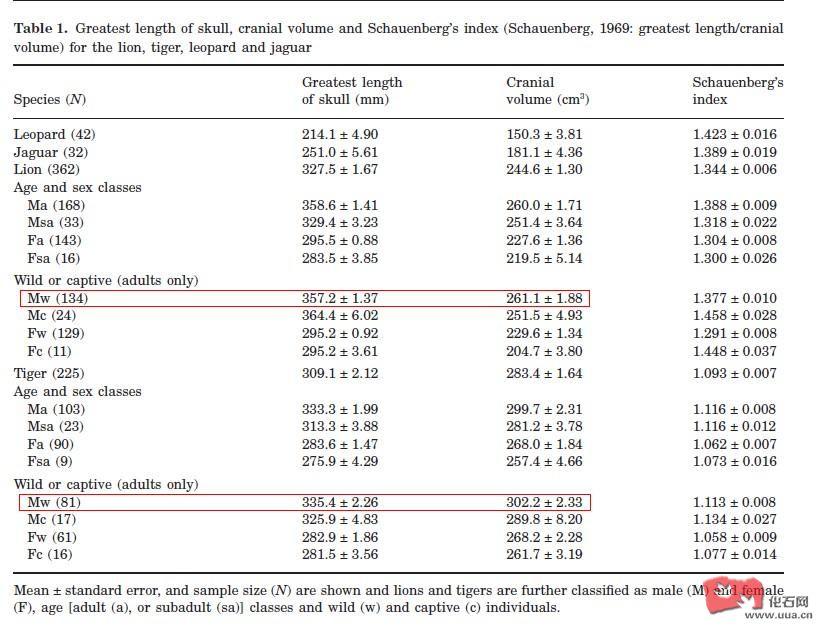
狮和虎的眼睛长度(Axis length),整体上狮的眼睛长度比虎要大些,有一个猞猁个体的眼睛长度居然比美洲虎要大,虽然说美洲虎取也不是什么大个体,估计是中美洲的小种群的雌性。狮子和虎的眼睛是否和他们的生活的
狮和虎的眼睛长度(Axis length),整体上狮的眼睛长度比虎要大些,有一个猞猁个体的眼睛长度居然比美洲虎要大,虽然说美洲虎取也不是什么大个体,估计是中美洲的小种群的雌性。
狮子和虎的眼睛是否和他们的生活的地方有关系,狮子在开阔地,而虎更多在半林地和长草地里面活动。里面狮的体重给的比虎小,虽然说应该和眼睛大小来源个体没什么关系,但是除非虎取得是幼年虎,否这狮眼睛Axis length比虎大,应该没有任何问题。其实这个问题看图也是比较明显的,可以看出差不多大的狮子的眼眶长度是比虎长一些
A是虎,C是狮子,不用理会测量尺,就这直接当成就这么大,狮的眼眶也长些,但是虎的吻部显得比狮子宽但是短的多,因此虎的前端咬力比狮子大,但是裂齿末端咬力相差就没那么大了。
狮子的眼睛是非常大的,即便是比狮子大的多的非洲郊熊的眼睛大小也远远不如狮子。头骨大小和狮子差不多的熊的眼睛就差更多了
现在看看肉食兽的咬力是这样计算的,肉食兽咬力最基本的计算方法是1991年JJ Thomason研创的三杠杆法(dry skull method),把头骨当成一个三节杠杆计算咬力,当然三杠杆的数学非常简单了,相对而言倒是Solid Beam Method稍微难些,国内不清楚,美国初中生都应该能做出来三节杠杆计算,高中毕业的时候做Solid beam也完全没问题,主要是难度是计算咬肌组和颞肌组切面的面积大小,这个需要高清晰的头骨照片或者头骨本身了。
步骤1
得到头骨照片并且计算面积,并且选定一种肌肉效率(举个例子,熊咬肌肉也许的可以选择类似人类手臂肌肉效率,或者直接用熊自己的,如果有研究了)
步骤2,3,采用公式计算咬力
最后得到咬力结果,需要注意的是这些力量只是半边头骨肌肉的力量,两边头骨肌肉一起咬的话力量大概是半边的1.7-2倍左右。下面是洞熊的咬力,可以看出雄性洞熊的咬力和一般的狮虎差不多,比大个体的狮虎略小一些,雌性洞熊则和较小的雌狮和雌虎差不多。
另外上面MF是咬肌,TF是颞肌,Ma和Ta分别是咬肌和颞肌的面积,我发现小问题,一般肉食目都是颞肌比咬肌大,颞肌几乎负责整个开口猛咬的过程,一直到完全嘴巴合拢咬实以后咬肌的作用才超过颞肌,但是洞熊居然是MF超过TF,Ma超过Ta,这个可能是要么颞肌大小被低估,因为颞肌是从里面穿出来的,而咬肌肉是外面的一层,看照片可能会低估了颞肌,因此洞熊的咬力只有大雌虎或者小雄狮很可能是个小地洞,其他作者的论文里面(日本专家组和Per Christiansen,Jan S. Adolfssen或者绝大部分的解剖学家都不会认为咬肌比颞肌的整体作用力大吧)。那雄洞熊的头骨比雄性狮虎大多了,咬力至少应该持平才对。
下面是一组更精确一些的MF和TF的咬力组合
第一是头骨本身的数据
下面的狮子是正常/(或者超大,假设SKL指颅基长)的雄兽,虎是个特大的雌虎或者小雄虎(或者普通偏小雄虎,假设SKL指颅基长),测量数据如上,另外吐槽下,Per Christiansen弄出一个basicarnial length,我一开始也不知道什么标准,一开始看表格的时候还以为他把宽度和长度高反了,后来去仔细查了原本的论文才发现是个新标准。Masseter是咬肌,temporal是颞肌,这里面颞肌全部超过咬肌肉,这个才是正常的肉食哺乳动物的头骨肌肉组,咬肌肉超过颞肌成丽齿兽了
这里面虎按比例算,咬力比狮子大,但是反之,只要狮子明显比虎大了,它咬力就能超过虎。但是看上面按比例算的话狮子的咬肌比虎发达,而虎的咬肌其实已经是非常发达的了,这个和狮子更多的用索喉窒息杀死较大的猎物有关系,咬肌的作用在嘴巴合拢的时候最大,虎的猎物一般较小,而虎习惯更多对中小猎物用较长的犬齿和强大的咬力直接咬断脊椎骨秒杀,因此虎按比例颞肌给的巨大力量猛咬,例如西虎主要的猎物野猪,索喉就不方便也许要先咬上了再杀,而西虎杀雌棕熊的话索喉可能就更不明智了
另外上面Cape的距离是上下犬齿之间的距离,不是说嘴巴之间的距离,上次发贴的太卡,没心情纠正。角度是开口角度。
此外上面还有狮虎的张嘴的度,显然嘴巴之后上下犬齿之间的距离10厘米左右,虎开口的度数略大于狮子,因为虎的吻部短,犬齿长,因此需要相对自己头骨长度更大开口上下距离来使用它的牙齿,上面张嘴巴度数不是最大的度数,但最大不应该超过表格上面度数太多。狮虎都根本没法张到一些垃圾科普上面说的110度,别说他俩那么大个冠状突,其实巴博虎科和毁灭刃齿虎那个形态的下颌骨和头骨的接连模式,长到110度也非常吃力的了,下面这个看似已经很夸张的张大嘴的巴博虎其实也就101度而已
也有老论文说虎的咬肌是最发达的,其实作者的意思是在该论文引用和研究的几个动物里面虎最发到,注意这个论文里面没有涉及狮子
下面这个虎其实是个苏虎,现在都可以算独立种,沪的咬肌肉分成了好几层呢,而小型猫科的咬肌就不如虎发达,另外我读了以后,那个表格的意思貌似是PT(Tubercle with developed process)的比T
早期1900-1940年有几个专家估测锯齿虎的头部和颈部的接连肌肉组要来之肩膀和头颈候补,按时其实另外和现代狮虎一样,起源于Altas,只是锯齿虎相对而言头骨的活动范围更大,颈部左右活动范围相对很大,1940早期的推测的锯齿虎攻击方式是靠颈部发力把头部当锤子用,下颌骨没有作用,而现在的估测这是头骨和下颌骨一起咬,下颌骨作用很大,其实剑齿虎亚科的下颌骨整体都比较强壮,别看冠状突比狮子小,下颌骨本身可不算弱。
下面是FRANÇOIS THERRIEN对剑齿型掠视哺乳动物的下颌骨的分析,对采用的方法和头骨不同,一般用的纠正过的Solid Beam方法,也就是空心杠杆法(实心杠杆法是用来算兽脚亚目,他们都是实心下颌骨,而哺乳动物和鳄鱼的下颌骨都是空心的),另外除了参靠下颌骨的形态,还要注意硬骨层的厚度,比如致命刃齿虎的下颌骨硬骨层的比例比狮子的要大。
计算的结果,居然好几个锯齿虎的下颌骨本身在裂齿处的强度,包括N型锯齿虎,锯齿虎sp.都超过了狮子,很大费氏巴博虎也超过了狮子,哪个狮子的下颌骨225mm,虽然不算特别大却不小了,另外巨型剑齿虎的下颌骨虽然没狮子大,但是239mm在巨型剑齿虎里面根本不算什么,西班牙的那个313mm的下颌骨肯定超过现代狮子(当然都没法和残暴狮比),另外虎在表格里面弱于狮子,其实没什么差别,因为虎用的个体比狮子长度小了8-9%。
下面是年代分布表,这个论文里面涉及的剑齿型也真够光的,居然袋剑齿虎里面的Thylacosmilus atrox和Nimravides都算近来了,Nimravides属也是像剑齿虎那样的,但是牙齿还是相对短一些,更接近云豹,它和当时的巨型犬熊亚科的Ischyrocyon都是当地的顶级捕食者,虽然后者地位明显更高些。
介绍一下solid Beam是的结果都代表些什么
Zx是指横向弯曲力量,X轴的上下弯曲,可以看上图就清楚了,Zy自然就是左右了,F=Zx/L,指的是对应颌骨安全指数的极限作用力。还有一个Zx/Zy的比例,如果大于1就是更宽,小于1就是更高。
如果是犬齿位置Zx/Zy > 1就是犬齿部位适合针对咬力造成的垂直压力,这样的颌骨适合猛咬猎物或者咬断骨头
如果是犬齿位置Zx/Zy =1就是犬齿部位同时适合针对猎物挣扎或者是控制猎物
如果是犬齿位置Zx/Zy < 1就是更适合控制猎物
另外美洲山狮是犬齿Zx/Zy=1.5,Zx/Zy比较大的话,在大猫里面往往就是速杀猎物,不需要用嘴控制挣扎的猎物,狼善于控制,云豹最善于猛咬,云豹的犬齿非常的长
另外这么看下面的表格,Zx/Zy已经说了,LOG(Zx/L)或者LOG(Zy/L)越小,相对颌骨抵抗对那个方面的作用力就越差,例如下图就是狮子最强(因为相对结构,以及绝对大小),云豹犬齿强过比自己大的猎豹,但是裂齿和臼齿不如猎豹,鬣狗相对坚固指数很大,臼齿强度居然超过了狮子,当然这个鬣狗相比那个狮子也够大的了,17.5cm vs. 22.5cm,但裂齿和犬齿不如狮子,另外鬣狗裂齿咬力其实只有狮子的一半都未必有,但是坚固程度可以和狮子匹敌,因为安全指数更多的是符合习性,有些习性例如啃骨头需要更大同样咬力下更大的安全指数。
猫科的咬力和咬力商,外加亲戚关系,咬力是残暴狮最大,比狮虎大多了,但咬力商残暴狮倒数,因为咬力商增加的指数是体重的0.67,一般越重的咬力商越小。
剑齿型的下颌骨强度介绍,指出一下,剑齿类按头骨算咬力商都很弱,按头骨算,但是按下颌骨很多个体算咬力商很强大,下颌骨都很强大,因此剑齿类的下颌骨没有用处,不是很靠普的说法。
残暴袋剑齿虎或者黑袋剑齿虎,犬齿对抗侧面力量不差,但是对抗垂直力量更强,上面那个个体蛮大的。袋剑齿虎体形和熊也有相似的地方,另外其的前肢粗壮,桡骨很大,几乎和胫骨差不多,袋剑齿虎是强大的伏击裂兽,这个大管家已经早说过了。袋剑虎的头骨除了枕骨部分略小,整体上接近雌虎的水平。有些人是袋剑齿虎会用头去撞它的猎物,感觉可能性不大
锯齿虎属,剑齿虎属,和刃齿虎属,注意致命刃齿虎的下颌骨的硬骨层比同样大小的狮子厚的多,所以下面的图片里面低估了刃齿虎的下颌骨强度,论文作者也注意到了,但是缺乏数据,没有纠正,如果头骨也是这样的话硬骨层比豹属的要厚,Woe算的finite element可能低估了头骨的强度,那么我按woe和PC结果引用的咬力就可能是地洞,虽然说目前没有找到刃齿虎的头骨硬骨层的研究,此外锯齿虎的下颌骨已经强度超过狮子很多了
巴博虎科的,下颌骨都非常的强,B.morris,下颌骨才15.39cm强度就可以和下颌骨20+厘米的虎匹敌了,巴博虎属的前肢力量杠杆效率和致命刃齿虎相等,费氏巴博虎什么略超过致命刃齿虎,而后肢也是巴博虎相对略强些,他们估测也是伏击猎手,控制了猎物以后速杀。
(在本文为化石网论坛精华帖,由网友鸿穹神皇上帝提供,仅供大家学习参考。编辑:刘琮滢)
2022-12-01