-

海河与沼泽之歌-前传
真鳄亚目(Eusuchia)在白垩纪时期兴起 , 目前没法确定牠们的先祖身份 , 在进化树上和真鳄亚目(Eusuchia)有关系的中真鳄类包括腔鳄科(Stomatosuchidae) , 副短吻鳄科(Paralligatoridae) , 伯尼斯鳄科(
真鳄亚目(Eusuchia)在白垩纪时期兴起 , 目前没法确定牠们的先祖身份 , 在进化树上和真鳄亚目(Eusuchia)有关系的中真鳄类包括腔鳄科(Stomatosuchidae) , 副短吻鳄科(Paralligatoridae) , 伯尼斯鳄科(Bernissartiidae) , 苏西鳄科(Susisuchidae) , 皱褶鳄属(Rugosuchus) , 吉尔克里斯特鳄属(Gilchristosuchus)及格伦罗斯鳄鱼(Glen Rose form)。 皱褶鳄属(Rugosuchus)与吉尔克里斯特鳄属(Gilchristosuchus)的归属位置未明 , 格伦罗斯鳄鱼(Glen Rose form)则是连一个种名都没有的化石样本。以上的鳄类的骨骼及鼻后孔都朝向真鳄类方向发展 , 比如鼻后孔后移至翼状骨范围内 , 椎骨从两凹椎朝向前凹椎方向转变 。
腔鳄科(Stomatosuchidae)是一个位置存在疑问的中真鳄类 , 也有专家认为腔鳄科(Stomatosuchidae)可能是真鳄类。腔鳄科(Stomatosuchidae)包括嘉峪鳄属(Chiayusuchus) , 腔鳄属(Stomatosuchus)及薄煎饼鳄属(Laganosuchus) , 牠们是滤食性鳄类。横带嘉峪鳄(Chiayusuchus cingulatus)被发现于甘肃省的晚白垩纪地层 , 仅只有不完整的牙齿化石被发现 , 横带嘉峪鳄(Chiayusuchus cingulatus)是一个疑难学名。
腔鳄属(Stomatosuchus)只有森诺曼阶(Cenomanian)埃及的无棘腔鳄(Stomatosuchus inermis)一种 , 无棘腔鳄(Stomatosuchus inermis)下颌长达2.1米 , 下颌长度是惊奇薄煎饼鳄(Laganosuchus thaumastos)的2.5倍 , 全长约10至12米 。无棘腔鳄(Stomatosuchus inermis)有长而平坦的鸭嘴型颅骨 , 下颌呈U字型 , 眶部在颅骨的背面 , 上颌骨前部分已具有多达30颗排列紧密的小齿 , 下颌可能具有类似鹈鹕的喉囊作滤食 。可惜唯一的无棘腔鳄(Stomatosuchus inermis)颅骨在第二次世界大战时被战火摧毁 。
无棘腔鳄(Stomatosuchus inermis)
薄煎饼鳄属(Laganosuchus)一共有两种 , 森诺曼阶(Cenomanian)尼日尼亚的惊奇薄煎饼鳄(Laganosuchus thaumastos)下颌长达0.84米 , 全长约4至6米 ; 森诺曼阶(Cenomanian)摩洛哥的西阿拉伯薄煎饼鳄(Laganosuchus maghrebensis)只有破碎的颌骨残骸被发现 , 西阿拉伯薄煎饼鳄(Laganosuchus maghrebensis)有相对狭窄的齿骨 , 齿槽也和惊奇薄煎饼鳄(Laganosuchus thaumastos)略有不同。腔鳄属(Stomatosuchus)及薄煎饼鳄属(Laganosuchus)的下颌骨冠突及颞上区之间有较短的内收肌 , 纤幼的下颌骨显示两者有很细小的咬力 , 两者可能在浅水中静止不动 , 仅仅张开嘴巴以守株待兔的方式捕捉猎物 。
西阿拉伯薄煎饼鳄(Laganosuchus maghrebensis)
惊奇薄煎饼鳄(Laganosuchus thaumastos)
副短吻鳄科(Paralligatoridae)是生活于晚白垩纪亚洲的中型鳄类 , 牠们的外观和短吻鳄类相似 , 具有宽阔的吻部 。 过去被认为有两个属 , 后来专家认为副短吻鳄属(Paralligator)只是沙漠鳄属(Shamosuchus)的同物异名 。沙漠鳄属(Shamosuchus)前颌骨具有小孔以便让下颌骨第四颗牙齿嵌入 , 显示沙漠鳄属(Shamosuchus)已具有短吻鳄亚科的覆牙合齿式 , 不过沙漠鳄属(Shamosuchus)没有下颌孔 。 沙漠鳄属(Shamosuchus)一些种类具有矮胖的后牙以压碎坚壳 , 尤其是龟类。沙漠鳄属(Shamosuchus)有较大的翼骨凸缘(pterygoid flange)及两行以上的皮内成骨 , 显示沙漠鳄属(Shamosuchus)的位置和真鳄类已相当接近 。
蒙古发现的沙漠鳄属未定种(Shamosuchus sp)
已知的沙漠鳄属(Shamosuchus)一共有十种 :
森诺曼阶(Cenomanian)乌兹别克的卡拉卡尔帕克沙漠鳄 (Shamosuchus karakalpakensis);
土仑阶(Turonian)或桑托阶(Santonian)蒙古的平额沙漠鳄(Shamosuchus gradilifrons) , 大沙漠鳄(Shamosuchus major)及乌勒盖沙漠鳄(Shamosuchus ulgicus) ;
平额沙漠鳄(Shamosuchus gradilifrons)
乌勒盖沙漠鳄(Shamosuchus ulgicus)
土仑阶(Turonian)或科尼亚克阶(Coniacian)乌兹别克的北方沙漠鳄(Shamosuchus borealis)及西方沙漠鳄 (Shamosuchus occidentalis) , 两者被认为是同物异名。
北方沙漠鳄(Shamosuchus borealis)
坎帕阶(Campanian)蒙古的德加多克塔沙漠鳄(Shamosuchus djadochtaensis) ;
德加多克塔沙漠鳄(Shamosuchus djadochtaensis)
早马斯特里赫特阶(Maastrichtian)蒙古的特氏沙漠鳄(Shamosuchus tersus) , 始祖沙漠鳄(Shamosuchus ancestralis)及乌兰沙漠鳄(Shamosuchus ulanicus);
特氏沙漠鳄(Shamosuchus tersus)
始祖沙漠鳄(Shamosuchus ancestralis)
乌兰沙漠鳄(Shamosuchus ulanicus)
松花江沙漠鳄(Shamosuchus sungaricus)被发现于吉林 , 化石属晚白垩纪 , 松花江沙漠鳄(Shamosuchus sungaricus)具有呈S形的股骨 , 股骨总长约162 mm 。 松花江沙漠鳄(Shamosuchus sungaricus)的背椎中枢长约33mm , 松花江沙漠鳄(Shamosuchus sungaricus)的背椎比始祖沙漠鳄(Shamosuchus ancestralis)的要大 。
松花江沙漠鳄(Shamosuchus sungaricus)
沙漠鳄属(Shamosuchus)种类颅骨一般长约11厘米至31厘米 , 最大的种类是大沙漠鳄(Shamosuchus major) , 大沙漠鳄(Shamosuchus major)颅长约51厘米 , 全长约4米多。
大沙漠鳄(Shamosuchus major)
众多沙漠鳄属(Shamosuchus)种类中 , 以晚白垩纪坎帕阶(Campanian)蒙古的德加多克塔沙漠鳄(Shamosuchus djadochtaensis)最能体现出中真鳄类及真鳄类之间的转变。在鳄类演化过程中 , 原鳄类的鼻后孔前端与上颌骨毗邻 , 中真鳄类的鼻后孔在上腭范围内 , 真鳄类的鼻后孔在翼状骨范围内 , 鼻后孔位置逐渐后移。此外真鳄类另一特征是具有前凹形(procoelous)脊椎 。德加多克塔沙漠鳄(Shamosuchus djadochtaensis)的次生腭在腭骨及翼状骨毗邻 , 德加多克塔沙漠鳄(Shamosuchus djadochtaensis)的鼻后孔位置在腭骨及翼状骨范围之间 , 这显示从中真鳄类至真鳄类的演化过程中 , 鼻后孔逐渐从腭骨范围后移至翼状骨。德加多克塔沙漠鳄(Shamosuchus djadochtaensis)同时具有前凹形的骶前椎及两凹的尾椎 , 这显示在真鳄类兴起前 , 在一些较优越的中真鳄类{或称新鳄类(Neosuchia)}已具有前凹形脊椎 , 真鳄类的起源要追溯至侏罗纪。
晚早白垩纪的农安皱褶鳄(Rugosuchus nonganensis)颅骨长约28厘米 , 属中型大小的鳄类。农安皱褶鳄(Rugosuchus nonganensis)与沙漠鳄属(Shamosuchus)同样有细小的眶下窗(suborbital fenestra) , 上腭同样有宽阔的内窗区 , 左右外鼻孔被鼻骨间隔分开 , 显示皱褶鳄属(Rugosuchus)和沙漠鳄属(Shamosuchus)有许多相似的特征。
农安皱褶鳄(Rugosuchus nonganensis)背部有两行以上的皮内成骨 , 皮内成骨没有前外侧突显示皱褶鳄属(Rugosuchus)和棱角鳞鳄科(Goniopholididae)关系疏远 , 并和真鳄类关系较近 。和吉尔克里斯特鳄属(Gilchristosuchus)不同的是 , 农安皱褶鳄(Rugosuchus nonganensis)的鼻后孔并不完全在翼骨范围之内 , 相对来说在进化树中 , 吉尔克里斯特鳄属(Gilchristosuchus)比皱褶鳄属(Rugosuchus)更接近真鳄类。
伯尼斯鳄属(Bernissartia)颅骨类似短吻鳄类 , 上颌骨及前上颌骨之间具有凹槽让下颌第四颗牙齿嵌入 , 伯尼斯鳄属(Bernissartia)有较钝的牙齿 , 牠们有典型水生鳄类的特征 。伯尼斯鳄属(Bernissartia)主模式种是弗氏伯尼斯鳄(Bernissartia fagesii) , 弗氏伯尼斯鳄(Bernissartia fagesii)生活于早白垩纪贝里亚阶(Berriasian)至阿普第阶(Aptian)法国及比利时 , 弗氏伯尼斯鳄(Bernissartia fagesii)全长约1米 , 在西班牙及英国也发现了伯尼斯鳄属未定种(Bernissartia)的化石 。
弗氏伯尼斯鳄(Bernissartia fagesii)
伯尼斯鳄属(Bernissartia) , 森林鳄科(Dyrosauridae) , 吉尔克里斯特鳄属(Gilchristosuchus)及格伦罗斯鳄鱼(Glen Rose form)的鼻后孔位置在翼骨范围之内 , 弗氏伯尼斯鳄(Bernissartia fagesii)与格伦罗斯鳄鱼(Glen Rose form)鼻后孔在眶下窗后缘 , 弗氏伯尼斯鳄(Bernissartia fagesii)第一尾椎是两凹椎 , 第二尾椎以后是前凹椎 , 显示两者已具有真鳄类的洐征 , 在德加多克塔沙漠鳄(Shamosuchus djadochtaensis)则没有这些洐征 。不过伯尼斯鳄属(Bernissartia)后眶及颅顶骨之间没有交汇 , 因此吉尔克里斯特鳄属(Gilchristosuchus)比伯尼斯鳄属(Bernissartia)要更接近真鳄类 , 不过伯尼斯鳄属(Bernissartia)的发现显示真鳄类可能源自欧洲 。
西班牙发现的早白垩纪伯尼斯鳄属(Bernissartia)颅骨化石
晚白垩纪桑托阶(Santonian)的特腭吉尔克里斯特鳄(Gilchristosuchus palatinus)在加拿大被发现 , 特腭吉尔克里斯特鳄(Gilchristosuchus palatinus)颅骨长约15厘米 , 特腭吉尔克里斯特鳄(Gilchristosuchus palatinus)的腭骨与鼻后孔交汇显示吉尔克里斯特鳄属(Gilchristosuchus)并不是真鳄类。特腭吉尔克里斯特鳄(Gilchristosuchus palatinus)的鼻后孔在翼骨范围之内 , 基蝶骨并不在脑壳的腹面 , 显示吉尔克里斯特鳄属(Gilchristosuchus)比森林鳄科(Dyrosauridae)要具更多洐征。吉尔克里斯特鳄属(Gilchristosuchus)后眶与颅顶骨之间的接触显示和真鳄类有密切的关连。
苏西鳄属(Susisuchus)应该是已知的和真鳄类关系最接近的中真鳄类 , 已知的有两种 。贝里亚阶(Berriasian)至巴列姆阶(Barremian)巴西的雅瓜里比苏西鳄(Susisuchus jaguaribensis)颅骨长约10厘米 , 全长约65厘米 , 雅瓜里比苏西鳄(Susisuchus jaguaribensis)颅骨具有额顶缝 , 上颞孔及颅盖骨背部有缝合显示和真鳄类有亲缘关系。
阿氏苏西鳄(Susisuchus anatoceps)生活于阿普第阶(Aptian)巴西 , 全长约50厘米 , 发现的阿氏苏西鳄(Susisuchus anatoceps)化石样本被认为是幼鳄。
在巴西桑塔纳组(Santana Formation)也发现了苏西鳄属相似种(cf. Susisuchus sp.)的肢体化石 , 化石属早白垩纪。
苏西鳄属(Susisuchus)有一个鸭嘴型的吻部 , 牙齿呈锥状 , 平坦的吻部能拨开水底中的泥沙以寻找鱼类 , 甲壳类及蠕虫等猎物。苏西鳄属(Susisuchus) 上颞孔后缘位置 , 鳞状骨及颅顶骨有交汇 , 显示苏西鳄属(Susisuchus)具有大多高等的中真鳄类(mesoeucrocodylians)的特征 , 不过苏西鳄属(Susisuchus)翼骨并入次生腭 , 苏西鳄属(Susisuchus)具有前凹椎 , 尾椎全是两凹椎 , 显示鳄类的椎骨从两凹椎朝向前凹椎方向转变(真鳄类具有前凹椎) 。 在分类树中, 苏西鳄属(Susisuchus)是相当接近真鳄类的祖先 。苏西鳄属(Susisuchus)的躯干能进行大辐度的侧屈以便在水中运动 , 但是苏西鳄属(Susisuchus)同时具有修长的前肢显示具有一定的陆栖性 , 苏西鳄属(Susisuchus)可能是水陆两栖型 , 修长的前肢能在陆上奔跑 , 也能在水底移动或拨开水草。
(本文为化石网论坛精华帖,由网友Sargon提供,仅供大家学习参考。编辑:常君滢)
2022-11-30
-

《怪兽复活》——之哺乳动物篇
根据最近半年热播的科普电视剧 远古巨兽复活和史前掠食动物系列 为蓝本文中介绍的动物全部采用拉丁学名;只要清楚是犬熊科 巨猪科,(豨科),鬣齿兽科就可以了重点介绍犬熊科,豨科则挑选几个较大的标本,鬣齿
根据最近半年热播的科普电视剧 远古巨兽复活和史前掠食动物系列 为蓝本
文中介绍的动物全部采用拉丁学名;只要清楚是犬熊科/巨猪科,(豨科),鬣齿兽科就可以了
重点介绍犬熊科,豨科则挑选几个较大的标本,鬣齿兽只是简单的参考一下,窃鹤和古巨蜥打打酱油。
注:下文中的动物翻译修订
残暴狮=北美似狮
巴氏虎属=巴博剑齿虎属
伟鬣兽属 =大鬣兽属
注2:以后我的帖子里面“南美巨型短面熊”放弃不用,都采用南美巨细齿熊
SectionI—Part I——犬熊简介
犬熊科最早出现在Eocene,在Oligocene晚期开始成为对其环境的生态产生举足轻重的作用,Miocene早中期属于鼎盛阶段,基本上在Miocene晚期灭绝,被猫科,恐犬,其他犬科和鬣狗型的掠食者取代,犬熊科中的大成员至少在很长一段时间内是他们生态环境中体型最大的动物,犬熊科本身分布很广,大到超过残暴狮,小的则和狼獾,欧亚猞猁等差不多,体型范围从小于等于5kg到超过200kg,甚至达到500kg以上,插手了几乎纯陆生肉食哺乳动物的所有生态地位
犬熊科大成员最著名的主要是(Amphicyon)犬熊属,属中又以新大陆的Amphicyon ingens和旧大陆的Amphicyon giganteus体型最大,但是除了犬熊属外还有至少还有两个属Ischyrocyon和Pseudocyon属的成员体型也可以达到犬熊属的水平,想比之下Ysengrinia,Pliocyon,Daphoenodon adilophontes则是现今虎豹的体型,daphoenus属基本上都是狼到狐狸之间,而paradaphoenus最小,估测也就4-5kg
犬熊科的进化趋势是越晚期的越大
中国,印度,北美,欧亚都有犬熊科的分布(虚线代表可能分布时间,实线代表已经确认)
犬熊 Part II——形态和体型
以上论文中根据现代棕熊参考得出Amphicyon ingens大个体上限水平体重600kg,可能略为高估,犬熊作为纯肉食哺乳动物,其体型可能要比棕熊稍微小些
实际上犬熊科的肢体介于猫科,犬科和熊科之间,大型犬500~600kg之间比较合理
图里面几条实线是现代猫科(felidae)肱骨形态增长趋势,而Fig-6虚线则是上犬(Epicyon)的趋势,越接近实线的和现代猫科越接近。
黑色圆圈是刃齿虎(smilodon),黑色方块是犬熊科(amphicyoninae),可以看出犬熊的肱骨和刃齿虎属比较相似
其余几个表格还有鬣牛兽科(oxyaeninae),鬣狗科(Hyaenidae ),袋犬科(borhyaenoidae),残暴狮/洞狮(panthera atrox/ spelaea)半熊犬属(hemicyon),上犬(Epicyon),猎猫科的伪剑齿虎属 (hoplophoneus),,已经《远古怪兽复活》节目里面的另外一个明星鬣齿兽科(hyaenodontinae), 锯齿虎属(homotherium)
Fig—7是肱骨长度(竖)对比肱骨中端直径(横),犬熊和猫科相对比较细长,犬熊甚至比等长的刃齿虎和残暴狮还稍微细一点,相对伪剑齿虎属,鬣牛兽科,袋犬科都是相对短粗型
Fig—6中肱骨肌肉对应的肩胛骨部分的长度[1](竖)对比肱骨全长(横),这次犬熊则是这段地区相当的长,代表它们的肱骨有相当大的力量,如果搞不清楚指什么部位,
-Deltoid Tuberosity就是肌肉附和的区域,犬熊科相对长肌肉也相当的多
顺便说一下,看Fig-4和Fig-5,半熊犬属相当的细,而锯齿虎其实蛮粗的,所以说锯齿虎比狮子瘦弱不成立,残暴狮的肱骨中端则是和等长的刃齿虎粗壮程度相等(但是残暴狮绝对长度大,自然绝对粗细也更粗)
[1]一开始看错了,以为是末梢,其实是侧面长度
犬熊 Part II——形态和体型
BORJA FIGUEIRIDO,JUANA. PÉREZ−CLAROS,ROBERTM.HUNT, JR.对犬熊的骨骼进行了更细致的描述,采用了除了肱骨和头骨全长之外的更多骨骼和指标
但首先搞清楚都是些什么指标
GLS:是最大头骨长度,CBL是基底长,ZW是颧宽,ML是下颌骨长度
犬熊 Part III——体重
北美犬熊属中最大的Amphicyon ingens的体重计算结果平均547 kg(GM),两个股骨公式的平均大概也是这个数,其余的犬熊属则基本上小一大截,大概和现代狮虎体型差不多
Ischyrocyon属比犬熊稍小(它计算结果虽然接近,但是那时因为都是高值,低值没有数据,所以对比对应值,明显比Amphicyon ingens小),Amphicyon ingens的其他数据也比欧亚大陆的同属成员更大,当然它也不一定是最大的犬熊亚科,因为犬熊亚科的Pseudocyon sp在Barstovian时期的年代较早的美国新墨西哥下颌骨标本(F:AM 49247)就有可能体型和最大的Amphicyon ingens相等的(按对应数据差不多大),平均大小则很可能更大,估测是同种的Pseudocyon sp年代稍晚的Clarendonian时期在北部(Nebraska)的头骨标本,(F:AM 25144)估测体重396 kg ,这个结果和另一位学则Van Valkenburgh 估测的F:AM 25144体重331 kg比较吻合,它和新墨西哥标本的体型差异有可能是性别或者地域/时空性,欧洲的Pseudocyon sansaniensis体重估测245kg,同样旧大陆的Amphicyon giganteus大概和Pseudocyon sansaniensis差不多。
注:Amphicyon ingens最大个体的体重定位其余的论文基本上确定在540~600kg之间,比起北美巨型短面熊最大个体的范围780~1000kg,和南美细齿巨熊的1400~2000kg来说相对要稳定地多。
犬熊 Part IV——头骨形态和大小(和其他巨熊比较)
犬熊科的大个体的头骨非常的长,Amphicyon ingens最大的一个标本基底长(BCL)就有520厘米,这个长度甚至超过了最大的育空短面熊和郊熊,哪怕是最大南美细齿巨熊(南美巨型短面熊)的头骨(颅全长560mm)基底长也未必能有那么大,所以犬熊很可能是纯陆生肉食目里面头骨基底长最长的了,颅基长比他大的也只能去肉食兽里面找安氏中兽属两种和伟鬣兽三位了(蒙古裂肉兽都差远了,蒙古鬣肉兽的全长才508mm),其他熊类的头骨长度除了南美细齿巨熊外即便是“枕基长”也几乎无法达这个标准,稍小一些的Ischyrocyon的最大个体的479mm的基底长仍旧把很多巨型肉食目甩开很大一截了
里面最大的非洲郊熊的颅基长也就465mm,虽然说墨西哥按下裂齿推测可能还有更大的郊熊属不确定种成员(但不一定肯定,材料太碎),但是即便等比例放大也就颅基长491mm,比起基底长520mm还差得很远呢
犬熊 Part IV——头骨形态和大小(和其他巨熊比较)
犬熊 Part V——犬熊的生态位
《远古怪兽复活》里面定位的犬熊是形态在大型猫科,熊科和犬科之间,那么它跟这些动物有些什么相同之处呢,它是不是真的纯肉食动物呢
有关这个问题,肢体的部分B. Sorkina对短面熊,郊熊和犬熊属以及Ischyrocyon都作了描述,并认为犬熊是纯肉食动物,而短面熊和郊熊至少追捕猎物的能力较弱。
首先可以看眼睛视觉和犬齿长度,视觉方面,郊熊agriotherium africanum 和巨型短面熊arcotodus simus都不如狮子,眼睛很小,犬齿也相对较短
犬熊相比之下,犬齿相对较长,视力虽然没有狮子好,却超过郊熊和短面熊(注意角度问题,如果犬熊头骨倾斜的角度和短面熊+狮子一样,眼睛还会更大)
短面熊介于猫科和熊科之间,甚至更接近猫,所以短面也未必不可能是基本肉食动物(肉摄取量占食物的90%),而犬熊相对短面熊更接近猫科豹属一些,半熊犬属和猫科豹属非常接近,几乎一样。
Amphicyon sp和熊科比较相似,而Amphicyon ingen和Ischyrocyon gidleyi完全在猫科豹属
和虎相比,Amphicyon/Ischyrocyon的桡骨的长度相对肱骨很长,而胫骨相对股骨较短,而狮子的胫骨占后肢相对长度比虎还要大,虽然狮和虎都是伏击猎手,狮子更多在平原空旷地带狩猎,需要跑得路更多(Stroganov,对比西虎的观测结果),而狼追赶猎物的距离远超过狮和虎,狼的后肢比例则胫骨占的比例更大,但是同样会长途追赶猎物的和班鬣狗则和犬雄有着相似的胫骨比例,但班鬣狗而桡骨的长度相对肱骨的比例则超过狼,根据桡骨,胫骨和肱骨判断这个估测犬熊是短距离伏击猎手,只会追赶猎物很短的时间
但是根据尺骨判断,犬熊属的尺骨相比虎,更接近狮子,但总体还是在狮虎之间,尺骨的杠杆效力超过狼,不过因为后肢比例的关系,还是不能狮虎那样像很快从卧倒(伏击需要的姿态)迅速站起加速追赶猎物,而Amphicyon ingen的脊椎骨的构造和豹属进行比较以后得出,Amphicyon ingen脊椎骨的构造可以让他增加跑步的速度和步伐的长度,却不能帮助其很快的加速。
据此判断,估测犬熊伏击能力比狮虎差
结论是犬熊会追它的猎物的时间长于狮虎,短于狼和鬣狗,而追击初速度则比狮虎慢。而和欧亚的大型犬熊属可能同时期的半熊犬属则是更加适应于追击猎物(尽管逊色于狼)
而犬熊的前肢强壮,可以压制挣扎的猎物,肱骨的deltopectoral crest很长,可以提供更多的肌肉,而且Amphicyon/Ischyrocyon的肱骨的关节更加灵活,但是犬熊属目前没有可伸缩的爪子。
犬熊的咬杀猎物的能力:精确度低于狮虎[1],因此像刃齿虎那样靠前肢压制猎物以后则靠硬咬肋骨或者颈部,结束战斗(这点和刃齿虎的精确攻击有别),犬熊是单独狩猎收,会选择和自己体型接近的猎物。
[1]注,大猫的胡子可以帮助它增加咬猎物的精确度(Turner and Anton 1997),犬熊的头骨类似狼,胡子没有那么发达。
(本文为化石网论坛精华帖,由网友鸿穹神皇上帝提供,仅供大家学习参考。编辑:常君滢)
2022-11-30
-
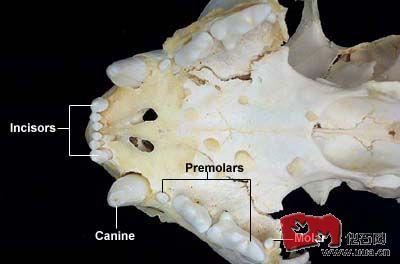
老虎头骨的详细描述
虎的头骨大而厚实 通常来说, 颅全长在 285~360MM之间 不同亚种间, 大小差异较大 所以, 285MM以下, 或360MM以上的, 也不少见 大概的范围是 雄虎:330~370mm,雌虎:270~310mm J H MAZAK 先生测量过
虎的头骨大而厚实。通常来说,颅全长在 285~360MM之间。 不同亚种间,大小差异较大。 所以,285MM以下,或360MM以上的,也不少见。 大概的范围是 雄虎:330~370mm,雌虎:270~310mm。 J.H.MAZAK 先生测量过的约300个虎头骨中,最大的有两个。 一个是采自乌苏里的北亚虎,颅全长为383MM,另一个是采自印度北部大吉岭的孟加拉虎,颅全长379~380MM之间。文献中记载的最大的虎头骨,颅全长超过了400MM。
虎颅形狭长,但与狮子相比,虎的头骨更呈拱状。 分为脑颅和面颅两部分,与狮类动物(包括拟狮和洞狮)相比,虎的面颅占的比例小一些。 虎的脑室部低而小,到后部有些变尖。 吻部较阔,眼窝较大。 颧弓非常宽大,极其向外延伸。 眶后突和颧骨眶突均钝而小。 二者距离甚远。眶间距较窄。 额鼻部高,除苏门达腊虎外,其它虎亚种的鼻骨狭长,个别虎鼻骨的延伸甚至可能超过上额骨的最后端。 额部中央较高,额骨和鼻骨缝的连接处呈深凹形。 上颌骨额突部向后不及鼻骨额突,枕部呈一锐角三角形。 矢状嵴和人字嵴发达。 尤其是大亚种的老年雄性个体,矢状嵴和人字嵴相当的突出。 腭部宽阔,听泡较宽大,似呈球状。下颌骨厚实,咬肌窝凹陷,冠状突甚高,关节髁较宽大。 其中"鼻骨额突通常要明显超过上颌骨额突,下颌骨的基缘几乎是平直的" 似乎是虎类独有的性状。
其实,按比例来说,虎头骨和豹头骨也很类似。 但是,除了上述差别外,按比例来说,虎的吻部要更宽,脑室部相对较小,乳状突相对较大,犬齿更加粗大挺拔,听泡按比例来说更小(这点不是很确定)。 而且,豹的头骨更容易呈现铁锈色,虎的好像更洁白一些。
虎的齿式为 I3/3,C1/1,P3/2,M1/1,总共30枚牙齿。 P4有相当发达的外前附尖,m1带有或多或少可分辨的下次尖,p4有一个发育较好的下次尖,一般比下前尖大; p3的下次尖和下前尖很小几近不能分辨。
虎的犬齿是现存猫科动物中最发达的。 平均来说,上犬齿长约55~65MM,一些北亚虎的上犬齿垂直高度甚至超过了75MM,最夸张的可达90MM。相比之下,狮的犬齿则相形见拙,平均长度仅仅50~60MM。 其它猫科动物更加无法与虎相提并论。
(本文为化石网论坛精华帖,由网友三千院凪提供,仅供大家学习参考。编辑:常君滢)
2022-11-30
-

《怪兽复活》——之巨型翼龙篇
巨型翼龙中包括了有史以来最大能飞行的动物,现在注意的介绍一下其中一些比较有名气的大个体下面是翼龙目之后的名字翻译翼龙目 PTEROSAURIA 61623 61623喙嘴翼龙亚目 Rhamphorhynchoidea (并系群) 61623巨型翼龙中包括了有史以来最大能飞行的动物,现在注意的介绍一下其中一些比较有名气的大个体
下面是翼龙目之后的名字翻译
翼龙目 PTEROSAURIA
喙嘴翼龙亚目 Rhamphorhynchoidea (并系群)
双型齿翼龙科 Dimorphodontidae
蛙嘴龙科 Anurognathidae
曲颌形翼龙科 Campylognathoididae
喙嘴翼龙科 Rhamphorhynchidae
翼手龙亚目 Pterodactyloidea
鸟掌翼龙超科 Ornithocheiroidea
帆翼龙科 Istiodactylidae
鸟掌翼龙科 Ornithocheiridae
无齿翼龙科 Pteranodontidae
夜翼龙科 Nyctosauridae
梳颌翼龙超科 Ctenochasmatoidea
鹅喙翼龙科 Gallodactylidae
翼手龙科 Pterodactylidae
梳颌翼龙科 Ctenochasmatidae
准噶尔翼龙超科 Dsungaripteroidea
德国翼龙科 Germanodactylidae
准噶尔翼龙科 Dsungaripteridae
神龙翼龙超科 Azhdarchoidea
朝阳翼龙科 Chaoyangopteridae
Lonchodectidae科
古神翼龙科 Tapejaridae
神龙翼龙科 Azhdarchidae
最大的实体化石翼龙基本上都是神翼龙科(或者翻译为阿兹特克翼龙科)的成员
其中材料研究最完善的是风神翼龙属,这个属包括了化石保存较完整,体型较小的“风神翼龙未未命名”Quetzalcoatlus sp.(也翻译为披羽蛇神翼龙,并以此为基础,根据残片推测出的北之诺氏风神翼龙;翼展最初估测的结果可达15米(Larson 1975)甚至18-20米(Larson 1975,Robert Bakkr 1986),也有估测为11米(langston 1981),13米(Henderson 2008)以及10-11米(Witton 2010),个人意见比较赞成11米的估测
风神翼龙的体重估测部分有低于100kg,应该基本上都不准确,可以无视,否则该动物的密度将低于30%,而最高估测544kg由于Henderson使用的模型不正确(采用的普通绘画版本,而不是正规的估测模型估测)而不取,目前最可靠的体重应该是Paul (1988),Maden(1994)Witton(2010)等人估测180-260kg之间。
以下是北之风神翼龙的1975年的新种报告,其实当时就有推测翼展11米(根据Pterodactylus antiquus/古翼手龙),但是早期材料偏向于大翼展,采用了用Pteranodon longiceps/长头无齿翼龙估测15.5米),风神翼龙的肱骨非常的粗壮,其实大型的翼龙的肱骨末梢(部分包括中段)都会比小型翼龙的更粗壮。而且很多大型翼龙在飞行涉及的部位有很多由于使用过度而导致的骨质增生,这些特别作为支持大型翼龙有飞行能力的证据之一。
北之风神翼龙和长颈鹿的比例,测量尺2米
上图:一些早期无齿翼龙属和大型风神翼龙的体重估测
其中有一些相当轻的体重并不是参考模型,而是用鸟类飞行原理推测出飞行的上限,并以此反推的,基本上低于150kg都没有什么参考价值。
风神翼龙的密度建议采用60-75%(高空长途飞行/靠气流浮行的鸟类密度~鸟类平均密度)
上图:翼展和体重的关系。另外文章里面提供了一些攻势
1.骨重量(kg)=0.065(体重)^1.071,鸟类
2.骨重量(kg)=0.061(体重)^1.09 ,哺乳类
3.体重(kg)= 0.55(翼展)^2.516,粗略估测,低估大型翼龙体重
4.体重(kg)= 0.681(翼展)^2.807,原始长尾型翼龙
5.体重(kg)= 0.519(翼展)^2.550,翼手龙
6.体重(kg)= 0.1833(翼展)^2.4823
7.体重(kg)= 0.206(翼展)^2.3329
8.体重(kg)= 0.1863(翼展)^2.4767
9.体重(kg)= 0.623(翼展)^2.346.特大翼龙公式(参考鸟)
0.体重(kg)= 0.5088(翼展)^3.0294.特大翼龙公式(参考蝙蝠)
研究显示至少中小型翼龙对应其体重的飞行能力并没有和现代动物相差过大。
可以看出一些较大的翼龙的翼负荷明显超过小型的翼龙,但是也不绝对,比如翼展(wingspan)2-2.5米这个级别的3种翼龙和3.7-4米级别的无齿翼龙类的差距也不是非常
2010年的估测(翼龙引用2008年的数据,注意2篇论文的第一作者是同一个人),并且给出了鸟类的参考,其中大贼鸥,游荡信天翁的翼负荷和一些翼龙明显有重叠了。 灰颈鹭鸨和史前的阿根廷巨鸟(表格上没有)的翼负荷很可能还超过北之风神翼龙
3D估重专家之一Henderson也对翼龙体重进行了估测,使用和Witton(楼上论文第一作者)一样的模型估测法,Henderson把估测过程列举出来了;我也稍微的极少一下,估测结果除了风神翼龙由于模型选择错误而卫星外,其余的估测应该都已经另外一些ie论文的作者人为是正确的,虽然有些和Witton的估重不一样。但是基本上在可靠范围内(Henderson的应该比靠公式推测更准确一些)
先列举一些做基模的鸟类和其密度数据,这些鸟类的密度相对较高,基本上都在75%以上(虽然说有些小部分特化的鸟只有60-75%)
这个才是正确的北之风神翼龙模型(根据同属小标本重建),上图不是按比例画的,胸腹部明显比Henderson的短,上图还有风神翼龙,无齿翼龙和信天翁的翼展结构比例图
基本上胸腹联合长度0.65米(不包括肩膀和骨盆),而Henderson的6.22米体长模型由于采用了不严谨的设定,放了一个胸腹部长度2.77倍的卫星结果造成那个544kg的天文数字,其实Henderson和Witton的修改模型重量为240kg。阿兹特克翼龙科的身体部分基本上和其他翼龙科差不多,只是由于其头部较大导致如果直接看体长,容易被误导。
下面就是错误的北之风神翼龙模型,重量544kg,身体太长,头相对太短
只需要注意的是风神翼龙实际体长只有5.1米,而体重240kg,但是其实去掉风神翼龙对表格的影响并不大
翼龙的胸肌相当的发达,肱骨也很强壮;值得注意的是2010年Witton突出的观点是大型翼龙的飞行能力推测不应该局限于鸟类和蝙蝠,比如说灰颈鹭鸨等大型鸟类需要长途助跑,起飞也颇为困难,但是翼展小的多得火鸡和孔雀却可以猛地串上去,而大型翼龙采用的也不一定是蝙蝠和鸟类一样的方式,例如翼龙更多会使用肱骨帮组弹跳(参考纪录片:恐龙无敌里面的镜头)以这种方式250kg的翼龙完全可以在立刻迅速腾空,而采用鸟类的助跑方式70-100kg的翼龙需要下坡路和正面大风甚至高空气流才可以起飞
翼龙的翼膜也可以提供超过鸟类30%的腾空能力,外加翼龙的肱骨相当的粗壮,强度参数商可达鸟类的2倍以上
相比之下翼龙股骨相当的弱,远不如鸟类,一般只有鸟类的一半,无齿翼龙等股骨甚至相对自己体型很短(还不如翼手龙),而阿兹特克翼龙科虽然按比例有比无齿翼龙科更长的股骨,然而强度依然很弱,这些都更进一步支持大型翼龙并没有放弃飞行而转向陆地,风神翼龙的理论飞行能力甚至不弱,可以靠自己的力量飞行1.5-2km,并且只要在这个距离内找到一个上升热气流,就可以进入省力的浮行模式了。
翼龙类的飞行模式
无齿翼龙科属于海洋气息型,依靠海洋气流飘行,捕食海面上的鱼类。这个符合无齿翼龙的发现地是古代海洋岸边的特点,相比之下在内陆环境里面出土的阿兹特克翼龙科则更偏向于热气流上升型和空中掠食者(例如科中较小的翼展3米的zhejianpoterus,当然一些无齿翼龙可能也是天空掠食者)风神翼龙应该是类似于安第斯神鹰那样的高空食腐动物。
美国德州出土的北之风神翼龙绝对不是同科中毫无悬念的最大者
在晚期MAA的Densuscedil-Ciula Formation(罗马尼亚)发现的Hatzegopteryx thambema/哈特兹怪兽翼龙的头部估测长度2.5-3米,翼展11-12米,至少和风神翼龙相等,其肱骨的末梢也比风神翼龙要长些(虽然Witton认为这是由于沉积变形导致的,但是即便如此哈特兹怪兽翼龙依然和风神翼龙并驾齐驱)
除此之外还有在美国东中北部分的菲利多菲亚发现的阿氏翼龙体型和很接近风神翼龙
其材料包括一块保存长度620厘米(估测全长720厘米)的颈椎骨,虽然原始材料已久丢失,但是仍然有测量数据和石膏模型保留了下来,假设720厘米是阿氏翼龙最长的颈椎骨(第5节),借此推测阿氏翼龙因该和北之风神翼龙差不多大或者略大于后者,估测翼展11-12米
当然一些材料宣称在墨西哥和南朝鲜,根据脚印估测有更大的翼龙
墨西哥的翼龙目前的论文还没有弄到,只知道脚印大概长度800mm,而朝鲜的脚印350mm,朝鲜的翼龙号称12米,实际长度根据无齿翼龙脚印推测应该只有8.2米左右,而墨西哥的翼龙应该理论值18米,但是考虑到大型翼龙翼展有缩短的趋势以及脚印形成过程中可能被放大,因此个人建议采纳13-14米的长度,大概体重300kg左右。当然不排除其实是兽脚亚目或者不能飞行的巨型翼龙的脚印。
最后需要指明BBC的鸟掌龙不过是个卫星而已,根本没有12米,至少没有指出任何12米的材料和鸟掌龙属有关系。最大的鸟掌龙类也不过6米而已。
朝鲜发现的翼龙脚
当地发现了528个恐龙脚印,443个翼龙脚印,以及上千个带蹼鸟类的脚印,底层是Uhangri Gormation的上层阶段,大概是8100-8500万年之间,年代为白垩纪晚期,在此之前以供依靠脚印建属的一共有3种翼龙,包括晚期侏罗纪怀俄明莫尔森阶Sundance Formation的Pteraichnus Stokesi,亚利桑那晚期侏罗纪莫尔森阶Carrizo Mountains的Pteraichnus Slatwashensis,此外还有Purbeckopus pentadactylus,来自早期自白垩纪的Purbeck Limestone formation。
在南朝鲜新发现翼龙被命名为Haenamichnus ichnogen,意思是海南郡翼龙
最大的一组脚印长度350mm
经过对比以后,根据形状和大小推测是一个新种。不是朝鲜已有的无齿翼龙或者Nyctosaurus无齿翼龙属的脚趾I-III相对纤细,而III制式略略的长于其余的两个脚趾,而南海郡翼龙的I-III长度相等,而且比较粗壮。
一般这个脚印的大小估测是最大的无齿翼龙(6-7米翼展)可以留下200-250mm的脚印,而阿兹特克翼龙科5-6米的翼展,一般留下200-220mm的脚印,而南朝鲜的大型翼龙脚印大部分都应该是阿兹特克翼龙科的,其中中等大小的脚印和中国的阿兹特克翼龙科Zhejiangopterus的很相似,据此推测最大的那个脚印应该是一个8.7-9.2米的翼龙,但是考虑到脚印可能有夸大,因此保守的推测8.4米左右。
以下是澳大利亚发现的翼龙,具体地点是澳大利亚西部,Miria Formation,发现地在古代是海边,年代是白垩纪末期MAA,有发现古两栖动物,当地出土过鹦鹉螺亚刚的Crimomia Tenuicostata, 珊瑚,以及少于牙齿,以期一些爬行类动物和一具兽脚亚目的肱骨,当然还有这个翼龙WAM 60.57的尺骨,根据对比了鸟掌龙属,无齿翼龙属,Santanadactylus,阿泽特克翼龙科准噶尔翼龙属等以后,认为尺骨和大型阿泽特克翼龙相对最接近,推测翼展3.6-4.9米,严格的说这个只能算是中型翼龙,但是澳大利亚本来翼龙就不算多,拿来打下酱油吧
这个翼龙的尺骨高密度骨壁浇薄,尤其是两端末梢更薄一些,末梢上面有2个明显的充气腔,末梢宽度53.5mm,保存长度134mm
中线宽度22.5 X 17.8mm
在法国的Merigon,晚期MAA,也发现了相当庞大的阿泽特克翼龙属,应该翼展有9米左右,或者接近9米,发现的化石是一块巨大的颈椎骨,形态比较接近风神翼龙和阿氏翼龙,颈椎骨保存长度460mm,估测完整长度550mm,比较小的风神翼龙属未命名种的480mm的颈椎骨大15%,作者还分析晚期翼龙科的分布其实也比较光,从美洲,欧洲,东亚到阿根廷和澳大利亚/新西兰,但是由于有限的挖掘力度,导致化信息缺乏,目前还不能直接下定论说白垩纪晚期大型翼龙种类多样化衰减。
首先介绍一个有意思的翼龙头骨,体型其实也不小
巴西,Crato Formation,白垩纪早期,Aptian,翼手龙亚目的Lacusovagus magnificens,意思是“巨大的浪迹江湖”的翼龙,中文名字暂时没有翻译,目前虽然可以确定是阿兹特克型态翼龙,但是还不能分入阿泽特克翼龙克,也许更加接近古神翼龙科
化石编号SMNK PAL 4325,推测头骨完整长度至少655mm,头骨似乎非常的中空,相对比较宽,最大宽度推测至少宽度190mm,在Crato Formation,第二大的翼龙是Tupandactylus imperator,头骨长度800mm,但是很大一部分是头饰,因此其下头骨从下颌骨尖端到方骨的长度其实只有390mm,该区域最大鸟掌龙的头骨长达420mm。因此毫无悬念的个化石是该化石点里面最大的翼龙,比原先最大的阿兹特克翼龙科成员的颌骨长了估测65%左右。
这个翼龙估测翼展4-5米左右,最保守的估测也应该略超4米,而且Lacusovagus magnificens作为阿兹特克翼龙属,拥有较长的肢体,站立起来的肩膀高度应该能达到1米左右,而一些Crato的大型的脚掌龙虽然翼展可能也能4-5米,但是他们站立起来高度只有Lacusovagus magnificens的一半
PS:作者还根据这个翼龙化石推测,一些朝阳翼龙可能成年以后也没有头冠,
巴西东北部的的Santana Formation也有翼龙化石,这个formation包括了aptian和Albian两个阶,下接Crato formation,上部分则是Romualdo Member,Romualdo Member还出土了激龙属的动物,这个Formation最有特色的翼龙恐怕要属古神翼龙科的Thalassodromeus Sethi了,意思就是驰海翼龙(sea runner),正模标本头骨长度1420mm(包括头饰),头骨本身的长度798mm,下颌骨推测长度710mm,保存长度635mm,编号DGM 1476-R,发现与于1983年。
这个翼龙和Lacusovagus magnificens一样有着非常大的鼻腔孔,以帮助减轻头骨重量
头冠上面发现了推测为血管的痕迹,血管组织相当的丰富,因此头冠的作用包括了降温或者充血变色炫耀等,驰海翼龙有着所有脊椎动物中最大的头冠,仅次于Tapejara imperator。当然驰海翼龙的冠也许有更多作用,例如头冠让头骨形成流线型的头部,加固了前颌骨,在于剪嘴鸥属做对比以后认为这个翼龙也许会利用头冠帮助涉入水中抓鱼,驰海翼龙有着相对阿兹特克形翼龙中最坚固的喙,结构和剪嘴鸥属也相当的接近,属于低飞涉水捕鱼者(当然作者并不认为它会潜水)。
当然南美洲最大翼龙远远不止5米的翼展,其实南美体型庞大的翼龙数量还真不少,下面就是一个列表,里面就有不少5-6米级
其中鸟掌龙科的Tropeognathus robustus和Tropeognathus mesembrance(现在已经改属名Coloborhynchus)拥有一个65cm的头骨和56cm的下颌骨,它俩就是BBC卫星鸟掌龙的原形。
当然真正大的材料是这个1993年发现的,发现于巴西西北部的Chapada Do Araripe,也是Romualdo Member/Santana Formation肱骨末梢,根据araripedactylus和Santanadactylus估测翼展8.6-9.3米,最保守推测(根据无齿翼龙属)8.2米一站,肯定是堂堂举行翼龙了,这个也是早期白垩纪最大的翼龙了,这显示出了晚期MAA和CAMP的阿兹特克翼龙属,早白垩纪也有大型翼龙。
现在出场的巨型无齿翼龙是无齿翼龙属的成员
Smoky Hill Chalk Member/Niobrara Fomration跨度400万年,从Coniacian一直到Campanian,翼龙方面最注重应该是无齿翼龙属,包括长头无齿翼龙(P. longiceps) 和史氏无齿翼龙(P. sternbergi),长头无齿翼龙的翼展可达5米,而史氏更是可达6-7米,甚至超过7米,一些观点是史氏翼龙其实是无效种,因为两者的骨骼几乎没有足够的形态差异,而头骨和体型的区别很可能是两性区分造成的。例如骨盆的产卵道,小型的“雌性”有着相对更大的产卵道,较大的雄性骨盆则反倒按比例产卵道较小。
无齿翼龙的肢体骨骼比较,可以参考北之风神翼龙的肱骨52厘米,翼展11米
还有头饰的差异。其中头冠的功能既然基本上限制与雄性,应该更多的是炫耀的作用,虽然说可能对捕鱼,气流调控,散热也有一些涌出
在北方的two Medicine Formation也有一些非常庞大的翼龙,保留一些骨骼残片,估测翼展7.5-9米,最初命名为一个新科Titanopteryiidae的成员,现在看应该是阿兹特克翼龙科的了
大型翼龙有的时候也会生病,例如有些情况下肱骨由于损伤,使用过度,感染或者其他原因
病症包括退化性关节炎,骨骼坏死,骨质增生,感染和骨骼伤口愈合以后的加固骨质,几乎所有的坏死之后都有骨骼增生现象可,主要的坏死发生在下颌骨和肢体骨骼,肢体和下颌骨的坏死也许是因为打架和受伤造成的,当然下颌骨坏死也许和食用腐烂的食物有关系。而肢体骨骼的退化性关节炎更多的是因为肢体走路和飞行的压力造成的。
化石已经证明一些情况下翼龙可以在退化关节炎发生以后任何正常的生活一段时间,股骨和指骨的损伤也有恢复的证据,虽然至少有一些菱齿翼龙和翼手龙因为翅膀受损而死。
还可以发一下拥有者奇特的头冠的古神翼龙科的图片,这些翼龙的头饰上可能有着非常显眼的色彩,写那些灰色是其实是薄膜
皇帝雷神翼龙等有着长达800mm的头骨,和一些灰色的软组织头饰,而另外一些如妖精翼龙属则有着实心的头饰。
(本文为化石网论坛精华帖,由网友鸿穹神皇上帝提供,仅供大家学习参考。编辑:常君滢)
2022-11-30
-
肉食恐龙复原比例图
侧面图片,从上到下:UCMP 137538,14 7米,14750-16500kg(UCMP 118742成年版和它差不多大)玫瑰马普龙非迪不控,14 7米,12750-13250kg (霸王龙MOR 1126和它差不多大)MOR 008,13 45米,12000-12750kg侧面图片,从上到下:
UCMP 137538,14.7米,14750-16500kg(UCMP 118742成年版和它差不多大)
玫瑰马普龙“非迪不控”,14.7米,12750-13250kg (霸王龙MOR 1126和它差不多大)
MOR 008,13.45米,12000-12750kg (霸王龙MOR 1152,UCMP 118742和它差不多大)
MUCPv-95,14.0米,10750-11500kg (撒哈拉鲨齿龙 新模和它一样大)
FMNH PR 2081,12.7米,10220kg
恐手龙正模,13.3米,9100kg (三角州奔龙和它差不多长,没它重)
霸王龙正模 12.0米,8700kg (君王蛮龙比它稍长,和它一样重)
南巨正模/纣魁龙 Paratype/伊迪加鲨齿龙最大个体 12.7-13米, 8400kg(南巨),8600kg(纣魁龙)
BHI 3033,12.0米,7650kg
MOR 555,11.6米,7250kg (三角州奔龙和它差不多重)
高棘龙NSCM 14353,11.84米,6300kg (玫瑰马普龙小型成年个体,撒哈拉鲨齿龙正模)
伊潘龙 6300kg
注:缺棘龙
总结:
按区域稳定种群
埃及棘龙的埃及和摩洛哥种群并列最大
晚期MAA的霸王龙区域平均第二大,南蒙大拿Garfield county区域霸王龙平均12-14吨(UCMP 118742 + UCMP 137538,其余的什么UCMP 137539这些没数据,都是碎的,估计其中成年个体和UCMP 118742差不多大),早期MAA的霸王龙和玫瑰马普龙并列第三大,那个最大的首领"非迪不控“可以算成年个体渐进水平的代表,理论上那些小马普中如果活的够久至少一半分能长到这个水平,摩洛哥的撒哈拉鲨齿龙第四大,渐进稳定在11吨左右,奥玛莎龙和南巨并列第四大,2个个体平均~13米,平均体重10吨(摩洛哥棘龙体平均不到12米)
如果所有的化石不论地点,不论时间短,不论分布光寡年龄大小,全部加除:
南巨整体平均第二大(略大于奥玛莎龙属),恐手龙,君王蛮龙第三大
这样算,发现化石多的,年龄段齐全的,跨度年月太广,种内差异大,小种群拖后腿 (孙德尔本斯孟虎一算进去,机械性按个体数量死算,孟虎按亚种平均没有瓦扎的病狮大)
原创的彩图,不同的肢体部分已经对过比例了,大象都是中恐的菊石画的,拿来作对比。
彩色较深的是移动肌部分,一半霸王龙的移动肌肉包括尾部辅助+肢体肌肉,肢体肌肉占体重25%,尾部肌肉占5%,整体占体重30%,腿部加上骨头重量大概占体重29%
最大的瑞氏古菱齿象 14000kg
最大的草原猛犸象 13000kg
最大的南方猛犸象 10000kg
最大的哥伦比亚猛犸象 10000kg
霸王龙
灰色(SUE) FMNH PR 2081 10250kg
蓝色(TERRA) LACM 23844 10050kg
黄色(Pecker) MOR 980 8900kg
绿色(Osborn)正模 CM 9380 8700kg
最大(Saber) UCMP 137538 15950kg
霸王龙从小到大,Scotty(这个体重很大,9500kg,但是头部比较小,东北加拿大霸王龙种群差异) 长度131cm,宽度90cm,BHI 3033(7850kg),长度141cm,宽度92厘米,FMNH PR 2081(10250kg),长度153cm,宽度97厘米,UCMP 137538(16000kg),长度171厘米,宽度119厘米,南巨MUCPv-CH1 长度180厘米,宽度60厘米,MUCPv-95,长度195厘米,宽度65厘米。
南巨取较大估测(Coria 1998),霸王龙取实体长度,UCMP 137538取等比例放大,南巨的下限估测是MUCPv-CH1 160厘米(宽56厘米),MUCPv-95,长度173厘米,宽度61厘米(Currie, Coria 2002, 2006)
霸王龙头骨还是兽脚亚目最大,长度估测是棘龙最大,南巨比棘龙小一点,下颌骨差5cm,头骨差10-15cm左右。也许棘龙和南巨一样大,但是霸王龙宽度太大,所以长度加宽度还是霸王龙,毫无悬念。
(本文为化石网论坛精华帖,由网友鸿穹神皇上帝提供,仅供大家学习参考。编辑:常君滢)
2022-11-30
-

从说更新世北美洲的哺乳动物
10000bc史前一万年,是一部古生物的盛宴。众多大家知道和不知道的古生物都将粉墨登场。因为有美洲剑齿虎登场,所以,描写的地点,一定是南北美洲。而南美洲是没有猛犸的(猛犸象居住在北美洲,欧亚大陆和非洲)所
10000bc<史前一万年>,是一部古生物的盛宴。众多大家知道和不知道的古生物都将粉墨登场。因为有美洲剑齿虎登场,所以,描写的地点,一定是南北美洲。而南美洲是没有猛犸的(猛犸象居住在北美洲,欧亚大陆和非洲)所以,电影描写的环境,就是更新世末的北美洲。
10000年前,正是风起云涌,冰川期结束的时候,动物们也是大量的灭绝。在北美洲,几乎60%的大型哺乳动物都灭绝了。在南美洲,广义的南方有蹄类,除了少数残余,剩下的也都灭绝了。在亚洲,猛犸,披毛犀等几乎全军覆没。华北动物群衰落,大型动物也在陆续灭绝中,华南动物群中,绝大多数大型动物全都终结了。欧洲几乎没有大的土著动物留下来。非洲偶蹄类也有极大损失,在大洋洲,几乎没剩下多少超过两米的动物。
这就是当时的时代背景。在这种前提下,我们的主人公登场了,他们究竟要面对什么样的危险呢。。。
随着一声鸣叫,一个战士消失了。一旁晃动的树木丛中,杀手露出了庐山真面目。它就是泰坦鸟。泰坦鸟身高近三米,统治着这个即将逝去的世界。虽然在影片中,它被称为"恐怖鸟",但是,大部分的恐怖鸟(Phorusrhacidae)早在一百五十万年前,就在南美洲消失了。电影中的它,是泰坦鸟。拥有巨大的双喙,可以肆无忌惮的攻击马,骆驼等各式各样的动物。虽然10000年前,马已经在北美大陆绝迹,泰坦鸟也走到了生命的尽头。但是,这并不影响泰坦鸟成为可怕的杀手。巨大的双喙,威力是惊人的。可以轻而易举的咬碎猎物的脑袋,还有粗壮有力的腿,也许真的能奔跑得很快呢。但是泰坦鸟的翅膀很小,也无法用翅膀攻击敌人。极其强大的嘴巴,发达的双腿,很小的几乎没什么用的翅膀。。。。。。这些属性不禁让人联想到另一个食肉动物之王----霸王龙。确实呢,也许,泰坦鸟就是大型食肉的恐龙的缩影呢。统治着这个阴暗的世界。但是,它用双喙爬树,还是有些夸张。因为它的体重实在是太惊人了。绝对超过200公斤。就算是双喙能一口咬碎人的脑袋,恐怕也无法承受自身的庞大重量。
最后的往往就是最厉害的,威名赫赫的美洲剑齿虎终于降临在荧屏上。虽然它的名字"剑齿虎"广为人知,但是,它并不是真正的剑齿虎。真正的剑齿虎属动物早就灭绝了。它是刃齿虎,也叫美洲剑齿虎。体型粗壮结实。恐武有力。但是体型还是没有电影里演的那么大。在沥青坑,出土了上千具化石,其中最大的,颅全长(脑袋长度),也只有378MM(南美洲的最大记录也并不大多少)。而目前有明确记录的最大的狮子则要超过400MM。所以,北美洲的美洲剑齿虎实际上比现代的狮子短小,但是,却更加粗壮强健。
值得一提的是,真正的剑齿虎,实际上是非常巨大的。亚洲的剑齿虎颅全长可以达到410MM,超过了现代的狮子和老虎,而非洲的剑齿虎则更巨大。甚至于可能是最大的猫科动物。但是,仍然没有电影里演的那么夸张。
美洲剑齿虎的上犬齿平均就有12厘米长,而现代西伯利亚虎,最大也不过9厘米。美洲剑齿虎可以把上,下颌张大到120度角,而现代的狮子和老虎一般也只有六,七十度,很难超过90度角。美洲剑齿虎的长牙,只有一个作用,就是割开对手的咽喉和血管,给对手带来致命打击。就像它的学名"致命的刃齿虎",而且,它的门齿也很粗大,可以帮助吃净猎物。美洲剑齿虎还有发达而灵活的颈部,可以准确的咬到敌人的咽喉,而不触碰大骨头,免得自己的牙齿被折断。还有粗壮的前肢。可以用来给别的动物很沉痛的打击。总的来说,它是非常可怕的动物。
其实,美洲剑齿虎是进化很杰出的动物。存在的时间很长,而且,其他动物,例如袋剑虎和猎猫类,也和它出现了几乎相同的剑齿。这就是所谓"趋同进化"。
没有物种能永远生存下去,大象,老虎,人类。。。有生就有死。每一个物种,有出现,就有灭绝,万物都是有始就有终。没有生命会永远存在,但我们还是要珍爱生命-自己的,其他生物的。
但是,刃齿虎究竟还是离我们远去。只剩下一具具骸骨,诉说着以前的八面威风。
大小姐写的,人类猎杀猛犸象
大约在13000年前,克罗维斯人来到美洲大陆,而与此同时,冰河时代末期的解冻期也随之来临。
一部分迁徙到现在美洲大陆的人建立了所谓的克罗维斯文化,该文化的遗址最初发现在一个叫克罗维斯的地方因而得名。这种文化在北美大陆各地都有发现,他们兴盛于13000多年前。猛犸象的庞大体型和厚厚的长毛无疑会成为人类食物和皮毛的目标。克罗维斯人以其大型石斧和尖锐石矛而闻名,这些武器和智人的战略对于从来没有遇到过这么强势对手的古骆驼、大地懒 、猛犸等大型哺乳动物来说,都是致命的。
考古学家的发现为这个假说提供了一些直接的证据。1932年在科罗拉多州的丹特地区发现了猛犸的骨骸和矛尖,而骨骼上有长矛留下的伤痕。此后陆续在其他考古遗址发现类似的情况。例如在亚利桑那州圣彼得山谷的雷娜农场,发现有十三具猛犸遗骸和石制的矛尖。这群猛犸象,看起来似乎是一个象群,聚集在一个水塘的时候全部遭到屠杀。
一名来自佛罗里达大学的研究员大卫·斯德曼,近来进行了一项科学研究。他从古巴和海地的群岛上搜集了一些化石,并用放射性碳测出了它们的年代。结果显示直到人类踏上美洲大陆的6000年后,古巴和海地群岛上才出现了人的足迹。而极富戏剧性的是,一种跟现代大象体形差不多大小的哺乳动物———西印度群岛地懒也在那个时候从那些群岛上消失了。
“如果气候变化是导致陆地树懒灭绝的罪魁祸首,那么岛上的树懒应该和美洲大陆上的树懒在同一时间消失,因为气候变暖在那个时候是全球性的。”斯德曼解释道。而且,如果气候是导致物种灭绝的最主要原因,那么很少一部分动物会受到影响,因为大多数植物和动物都能适应气候的变化。然而事实却是相反,大陆上的树懒早在千年以前就灭亡了,而岛上的树懒却在人类到达之后相继消失,难道这是巧合吗?
这也解释了为什么冰河时代,北美许多地区四分之三以上的大型哺乳动物,包括猛犸、乳齿象、美洲剑齿虎以及巨型短面熊等物种,在短短的几千年间就全部灭绝了。
大卫·斯德曼也认为,气候变化在巨兽与人类统治权力更替的过程中仍然起着重要作用。它使得那些大型动物体质变差,从而更难抵挡来自人类的攻击,最终被挥舞着武器的人类永远赶出了这个世界。不知道大卫的解释能否最终终结这些大型动物灭亡原因的争论?
回复人:三千院凪
中文名 泰坦鸟
学名 Titanis
释义 巨大的鸟
分类 鸟纲 今鸟亚纲 今颌总目 鹤形目 恐鹤科 恐鹤亚科
体高 可达2。5米
体重 可达150公斤
分布 美国佛罗里达州和德克萨斯州
食性 肉食
时代 上新世
命名者 Waller 1963
泰坦鸟属于鹤形目(Gruiformes)恐鹤科(Phorusrhacidae)恐鹤亚科(Phorusrhacinae)的巨大肉食鸟类,是恐鹤科中最后的成员,泰坦鸟属下只有一种,称为巨大泰坦鸟(T。 walleri),由Benjamin I。 Waller在1963年命名。泰坦鸟体高可达2。5米,体重可达150公斤,是最大的恐鹤亚科肉食鸟类。和其他的恐鹤科一样具有巨大的喙,而且它的退化双翼不可能用来飞翔,但是进化出了一对用来抓住猎物的爪,这种尖钩爪可以轻松撕碎猎物。泰坦鸟的中趾则巨大,可以踢抓猎物。泰坦鸟生活在上新世的北美洲,分布在佛罗里达州和德克萨斯州,一般认为恐鹤科在北美洲和南美洲连接的时候因为更进步的肉食动物的竞争而灭绝了,但是泰坦鸟却反向进入北美洲,直到180万年前才灭绝,但是1995年在德克萨斯州一个更新世沉积中发现了泰坦鸟化石,显示它们是直到15000年前才灭绝的,这就是说早期的美洲人类很可能不得不面对这些可怕的肉食鸟类。
泰坦鸟。jpg (43。72 KB, 下载次数: 81)
中文名 美洲剑齿虎,刃齿虎
拉丁文学名 Smilodon
释义 刃齿
分类 哺乳纲 真兽亚纲 食肉目 猫科 剑齿虎亚科 刃齿虎族
体长 1。2-2。5米
体重 50-300公斤
分布 北美洲和南美洲
食性 肉食
时代 上新世-全新世(300万-1万年前)
命名者 Plieninger 1846
说起致命刃齿虎(Smilodon fatalis),它应该也是和恐狼,郊浪一样的机会主义者。属于剑齿虎亚科,刃齿虎属。Smilodon由于媒体的宣传而大名鼎鼎。其实,它并不是真正的剑齿虎。而应该叫"刃齿虎"或"美洲剑齿虎"更合适。真正的剑齿虎应该就是短剑虎(Machairodus)。
现代发现的刃齿虎属,有三种。生活在从上新世到更新世的北美洲和南美洲。分别为:纤细刃齿虎,致命刃齿虎,一般刃齿虎。在沥青坑,只发现了致命刃齿虎。其它的,纤细刃齿虎和豹子差不多大。一般刃齿虎比致命刃齿虎更大。但是体型还是没有电影里演的那么大。在沥青坑,出土了上千具化石,其中最大的,颅全长(脑袋长度),也只有378MM(南美洲的最大记录也并不大多少)。而目前有明确记录的最大的狮子则要超过400MM。所以,北美洲的美洲剑齿虎实际上比现代的狮子短小,但是,却更加粗壮强健。
值得一提的是,真正的剑齿虎,实际上是非常巨大的。亚洲的剑齿虎颅全长可以达到410MM,超过了现代的狮子和老虎,而非洲的剑齿虎则更巨大。甚至于可能是最大的猫科动物。但是,仍然没有电影里演的那么夸张。
致命刃齿虎的雄性,颅全长能超过350MM。几乎和现代的几个大型虎亚种或者狮类相当。沥青坑中发现了有伤残痕迹后康复的致命刃齿虎,所以,有人说它们食腐或群居。其实都是不合理的。首先,致命刃齿虎的上犬齿太大,食物利用率不会高。让它啃骨头是强人所难。其次,现代的一些群居动物,受伤后会离开群体,避免成为大家的累赘。而且,独居动物受伤后,也有康复的例子。
最近的研究表示,致命刃齿虎的咬力可能很小。这个结果好像和异特龙(ALLOSAURUS)类似
中文名 猛犸
拉丁文学名 Mammuthus
释义 地下潜伏的动物
分类 哺乳纲 真兽亚纲 长鼻目 真象亚目 象科 象亚科
体长 4-6米
肩高 2。5-5米
体重 4-12吨(平均4-8吨)
分布 非洲,欧洲,亚洲和北美洲
食性 草食
时代 上新世-全新世(480万-4500年前)
命名者 Brookes 1828
猛犸是最著名的灭绝动物,它拥有长而弯曲的象牙,而且虽然应该不是所有的猛犸都有长毛,但是北方的种类经常长有长毛,所以也叫长毛象。它们生活在480万年前的上新世到4500年前的全新世,并且广泛分布在非洲,欧洲亚洲和北美洲各地。英文猛犸(Mammoth)一词来自俄文mammut,而俄文来自西伯利亚民族语言中的“地下潜伏的动物”,但是西伯利亚民族没并不了解猛犸,他们从深达数百米的冻土中挖出猛犸的尸体,认为猛犸是生活在土里的,见到光就死的巨兽,因为他们发现的猛犸都已死亡并半埋在土中。
尽管猛犸起源于非洲祖先,但是实际上和现代亚洲大象关系密切,而且其实亚洲象属也是在非洲起源的。猛犸和亚洲象的共同祖先和非洲象的分化发生在持续干旱大约的600到730万年前的中新世晚期的东非,亚洲象和猛犸的分化发生在大约50万年后。猛犸的最早种类是非洲猛犸,最初出现在大约480万年前的上新纪的北非,化石在乍得,利比亚,摩洛哥和突尼斯都有发现。另一个最古老的种类是南非和肯尼亚的南非猛犸,大约生活在400万年前。非洲的猛犸后来迁徙到了欧洲并进化出了南方猛犸,南方猛犸又进入了亚洲还通过了白令陆桥到达了北美洲。在中更新世温暖的气候明显恶化了,因此欧洲,亚洲和北美洲的大草原和森林变成了不肥沃的干草原,因此南方猛犸和帝王猛犸被更适应寒冷气候的可能起源于170万年前的华北(魏光飚 2003)的草原猛犸取代了。真猛犸出现在大约30万年前,真猛犸更好能应付冰河时期的极端寒冷,最终在晚更新世取代了草原猛犸。真猛玛是猛犸属非常成功的一个种,它们在从西班牙到北美洲有大量种群,俄罗斯科学院的研究人员估计在冰期期间,西伯利亚的部分地区的真猛犸平均密度甚至可以达到每一百平方公里六十头。
大部分猛玛都在最后一个冰时期灭绝了,但是少部分在圣保罗岛和阿拉斯加存活到了公元前6000年,而俄罗斯朗格岛的猛玛直到公元前2000年才灭绝。一般认为猛玛是因为人类的疯狂捕杀灭绝的,另一种理论认为猛玛也许是人类的传染病和气候变化的受害者。气候变化和人类的捕杀的组合大概是最可能的一个原因。新的研究表明,即使人类的狩猎可能不是猛玛灭绝的主要原因,但是人类的狩猎可能也是一个重要原因,因为早在石器时代人类就开始捕食猛玛。矮小的猛玛之所以能生存在的朗格尔岛,是因为距离有人居住的大陆非常的遥远,相似的变矮小发生了与在加利福尼亚的侏儒猛玛。
一种常见的误会是认为猛玛大于现代的非洲大象,导致了猛玛作为“非常大”的形容词使用的错误。尽管猛玛中最大最著名的种类,加利福尼亚的帝王猛犸和北美洲的哥伦比亚猛犸肩高肩高一般3米多,甚至超过5米,通常大约有8吨重,但是特别大的公象也许可以超出了12吨重,超过了两头非洲大象的体重,这些巨大的动物是仅次于几种巨犀和恐象的陆地动物。但是猛犸的大多数种类包括最著名的长毛象真猛犸只有一头现代亚洲象那么大,而矮小的猛犸种类有在加利福尼亚州的侏儒猛犸和矮小的真猛犸亚种生活在俄罗斯西伯利亚北部,北极圈之内的朗格岛。
根据猛玛的近亲现代大象估计猛玛的社会结构大概也类似非洲象和亚洲象,一个老年母象带领和自己有关系的母象和小象组成象群,公象在性成熟以后单独生活或组成松散的小群。猛犸头骨短高,正面呈弓形,侧面视顶部成圆顶三角形,额顶部隆起呈圆形或比较平坦,额部下凹,枕脊凸很高。臼齿宽大,釉质层薄褶皱细小规则,厚1。5到2毫米,各齿板间的齿质层和水泥质层厚度大致相等。臼齿齿脊频率18到27个,有些可能略少,最多可以有30个。齿板整齐紧密匀整,数目多。猛犸的门齿很长并强烈旋卷弯曲,最长的可达4。8米。而真猛犸背部长有一层绒毛和长毛,真猛犸背部的毛发最长可达50厘米,皮下脂肪厚达9厘米,真猛犸的头颈部分还有高大的驼峰可以储存大量的脂肪。
现在陈列于圣彼德堡动物博物馆的“Berjosowka”是1900年在别列索夫卡发现的猛犸尸体,胃里有山榛,白桦,艾蒿,大看麦娘,百里香,阿尔卑斯罂粟,鹿蹄叶,毛莨,地榆,花苔,落叶松和开花的菊科植物,显示死亡时间是夏末或初秋,并没有冻土地带的代表植物,其中只有大看麦娘和阿尔卑斯罂粟可以在冻土地带生长,而白桦,花苔和落叶松现在在别列索夫卡没有,生长在靠南一些的地方,而别列索夫卡现在的的地面上只有少量的积雪了,植物主要是针叶林。
另一个在雅库次克山德林河中游右岸发现,胃已经破碎,肠子里百分之九十是草本植物:禾本科和香蒲科的叶和茎的碎屑,还有柳树,赤杨等的嫩枝叶禾一些青苔,因为食物里有未成熟的种子,说明死亡时间在初夏。
1977年在俄罗斯发现的“迪玛”是最著名的猛犸尸体,它是冰川压蹩了的幼年公猛犸。俄罗斯科学院的研究人员认为不可能克隆猛犸,然而保存良好的猛犸尸体对研究猛犸是有用的。美国的拉布雷亚沥青坑都是著名的猛玛产地。
乳齿象和猛犸象模样很像,也很大,对二者牙齿的研究说明,猛犸象爱吃草,乳齿象爱吃树叶。
关于区别
1,当地发现的美洲乳齿象(Mammut americanum)属于短颌乳齿象,猛犸属于真象。
2,在上门齿,就是象牙上,猛犸的显得更长,而且扭曲的极其厉害。
3,在臼齿上,美洲乳齿象的牙齿是脊形齿,适合吃叶子。猛犸的是齿板非常密集的臼齿,适合吃草。
4,猛犸的头部高耸,乳齿象较低。
5,在大的骨头之类的,例如前肢,乳齿象的肱骨和肩胛骨按比例来说,是比猛犸象小一些的。
6,在骨盆上也有一些区别,但是这个也可能是性别不同造成的,所以我就不赘述了。
【转贴】史前一万年
史前一万年 10,000 BC
基本资料
制片:Roland Emmerich
Mark Gordon
Michael Wimer
导演:Roland Emmerich
编剧:Roland Emmerich
Harald Kloser
摄影:Ueli Steiger
制作设计:Jean-Vincent Puzos
配乐:Harald Kloser
Thomas Wanker
演员:Steven Strait 。。。 D'Leh
Camilla Belle 。。。 Evolet
Cliff Curtis 。。。 Tic-Tic
Omar Sharif 。。。 Old Baku
Nathanael Baring 。。。 Baku
Mona Hammond 。。。 Old Mother
简介
在一个遥远偏僻的高山部落,年轻的猎人达雷(史蒂文&#8226;史崔特饰演)找到了心上人 – 美丽的伊芙乐(卡米拉&#8226;贝尔饰演)。但神秘外族入侵,村庄突遭洗劫,心爱的恋人被抢走,于是达雷毅然决定率领小簇族人踏上漫漫追踪之路。当他们第一次来到未知的大地,发现了文明的存在,人类的生存方式远远超出了他们的想象。一路颠簸中,不断有其他部落同样遭受灭顶之灾的人群加入,不知不觉中达雷的追踪小队壮大成了一支军队。命运的驱使,让毫无作战经验的勇士们必须面对史前猛兽的袭击并不断与恶劣的大自然作殊死搏斗。在即将结束他们英勇顽强地长途跋涉的时候,他们偶然发现了失落的文明,大金字塔飞奔入云的壮观景象让他们最终领悟到他们的终极命运被一个无法想象的帝国掌握。他们必须抵抗帝国的统治者,一个自称神灵的暴君。达雷在这场腥风血雨的洗礼中终于明白:他的使命不仅仅在于营救伊芙乐,而是拯救整个人类的文明。
史前时代的大冒险传奇
《史前一万年》是全球卖座大导演罗兰艾默瑞奇(《ID4:星际终结者》、《明天过後》)的最新特效钜作。本片描述一趟伟大神秘的探索旅程,进入充满预言及传奇的远古神话年代,当时的世界由神祇及精灵统治,而巨大的史前动物则摇撼著地球。
在偏远的一个山间部落中,年轻的猎人达力遇上他梦寐以求的美丽女子伊芙乐。然而,一批邪恶的蛮族兵团突然袭击达力的村庄,并劫掠了伊芙乐。为了拯救深爱的女人,达力率领一群猎人踏遍天涯海角试图找到那些蛮族兵团。在追踪的过程当中,这群猎人进入了许多未知之地,并得知其他文化的存在,这些意想不到的发现令他们大开眼界,而且每次到达一个新世界,当地苦於蛮族兵团侵害的部族便会加入他们,於是达力原本的猎人小队逐渐壮大成为雄伟的部落联军。
在命运的逼迫下,达力率领的这群战士必须跟猛兽及掠夺者对抗,同时还要忍受最严厉的外在环境。在英勇旅程接近尾声之际,他们竟然寻获一个失落的文明世界,并瞭解到他们的最终命运操之於一个超乎想像的帝国,那里的金字塔几乎深入了天际。
此刻,他们必须击溃一个奴役民众的专横神祇,而达力也终於领悟到,他的使命不只是救出伊芙乐,还有挽救一切的文明……
一名远古英雄的旅程
《史前一万年》导演罗兰艾默瑞奇(Roland Emmerich)拍过许多令人留下深刻印象的大片,主题从星际大战到全球灾难都有,执导过的卖座强片包括《ID4:星际终结者》及《明天过後》。这次,他把镜头移到远古的过去,打造「史前一万年」的世界。这部电影可说是这位导演最有野心、最具挑战性的作品。
这个故事关於一位出身於偏远部落的猎人英雄,勇敢对抗一个巨大的帝国。为了创造这出神话故事,罗兰艾默瑞奇努力打破电影媒介的限制,带领观众经历一趟前所未有的冒险历程。他表示:「我一直很喜爱传统的说故事方式,就是大家围著营火,上一代为下一代讲述永恒的故事。当故事的主题是早期人类时,说故事者就大有机会编造一个全能英雄的传奇寓言。我拍摄这部电影,是想让观众彷佛进入一个不同世界,里头的景观和氛围都是前所未见的。」
为了带领观众踏上这趟不同时空的冒险旅程,罗兰艾默瑞奇率领其演员与制作团队远赴世界的另一端。拍摄地点从冬日严寒的纽西兰、湿热的南非开普敦,到乾燥的非洲那米比亚国沙漠。
制作人麦可威玛(Michael Wimer)表示:「罗兰艾默瑞奇总是想制作出具有原创性的东西,但电影中想创造出新意可不是一件容易的事。以各个层面来说,这样的目标都具有极大的挑战。事实上,罗兰艾默瑞奇自己也说过这部电影是他拍过最困难的作品,不过对於像他这样的导演,挑战性反而是向上的动力。」
哈洛德克罗瑟(Harald Kloser)跟罗兰艾默瑞奇合作为这部电影编剧,他表示:「《史前一万年》的背景当中,神秘性及精灵界是组成生物世界的主要部份。我跟罗兰艾默瑞奇从未想过把《史前一万年》这样的题材拍成一部纪录片。相反地,我们要创造出一个伟大的人类冒险旅程,史前的人类必须面对一切他们无法解释的力量。我们都喜欢向极限挑战。」
制作人马克高登(Mark Gordon)这次是第三度与罗兰艾默瑞奇合作,他表示:「罗兰艾默瑞奇是那种绝不愿意拍摄重覆题材的导演。他的想像力让他能够神游到一般人无法到达的境界。这个故事合他胃口,而且他的视觉表现又极为震撼,所以他是执导这部电影的最佳人选。」
幕后制作:
穿梭于洲际之间
创造史前一万年的世界
艾默里克与自己的幕后创作团队为本片创造出了一个原始荒芜的史前世界,观众无法想象也从未有过如此逼真的感同身受。尽管影片没有说明故事发生的特定地点,但艾默里克一直觉得是在非洲。“那里是人类的起源,”他说,“但因为我们的故事,它就变成了我们自己想要的非洲。”影片的拍摄地点主要在新西兰周边的一些颇具特色的地方,并在非洲选择了一些故事情节需要的地点。包括开普敦,南非以及纳米比亚的月色。
影片原本计划只在新西兰拍摄数日,但在开拍前六周的一次直升机空中选景时,艾默里克被这片迷人的“伊甸园”深深地吸引住了。 “我们花了一整个上午在空中选址,刚回到酒店还没坐下就收到艾默里克的紧急留言让我们立刻回到直升机上,说他发现了更棒的东西。 制作人维摩回忆说:“当时我都已经准备劝说艾默里克,开机时间就在眼前了,不能再随意更换拍摄地点了。但还没机会开口,我就已经重登直升机,当飞机上升到某一高度,艾默里克痴迷的那一片天地突然出现在了我的眼前,和剧本里的情节描述一模一样,感觉剧本就是根据这里描写的,我们别无选择,就把拍摄地点定在了那里!”
冰天雪地、乌山黑林的景色会和南非热带丛林的郁郁葱葱形成夺人心魄的强烈对比,影片的中间部分依靠的就是这种自然的背景天幕。 纳米比亚地域燃烧般炙热的橙色和红色也为影片近三分之一的情节提供了巧夺天工的拍摄条件。 正因为有了这些场景,新西兰的“白色和黑色”场景就越发显得魅力难挡。所以摄制组为了能如愿拍到最好的景色,向新西兰地区千变万化的天气做了太多的妥协和让步,根据随时而来的雾雪风暴或立即万里无云的反常变化作适时地拍摄进度调整。
维摩说:“我们特别希望通过掌握当地地形的特色体会我们剧中角色在当时那个年代是如何艰难的生活着的,但同时,在如此广博的天地中生活,又是件多么美妙的事情。 正因如此,我们必须在那里拍摄,感觉特别的宏伟,美得不真实。”
拍摄地点风景如画,也为拍摄组提出了各种要求极高的保护措施。“为了不留下过多过重的痕迹,我们使用了四轮车调动各种拍摄器材,拍摄过程中尽量采用超轻量级小车,这样车轮碾过草地也不至于留下痕迹。”新西兰拍摄场地主管杰拉德&#8226;康农说。 “我们还大量依靠直升机来回搬运用于搭建亚高族部落的道具和布景。”
怀奥鲁雪场位于离新西兰瓦纳卡镇(世界著名的试车基地)不远处的南部岛屿上,海拔5000英尺,为本片的拍摄提供了五个重要场景。包括亚高族部落的村庄,巴库的石头,杀戮现场和牧草地。影片几乎有三分之一的部分在那里拍摄完成。新西兰境内其他的拍摄地点包括艾斯派林山国家公园和普奔水坝。
对艾默里克来说,雪场与他脑海中想象的影像不谋而合。“这种地方能让你从高处随意掌握镜头的角度,感觉就象漫游月球,你拍到的东西就象在月球表面看到的,”他兴奋地说道:“给你很多你想象中远古时代的感觉。我们剧中的人物在旅途中展开他们的故事,我们需要宏大的场景将现在的世界转变成他们旅途中进入的世界。所以环境的千变万化是非常有用的。”
在开始基本场景的拍摄前,制作组邀请了 Ngāi Tahu (南部区域毛利族的酋长) 去实地考察一处即将用以举办传统毛利族庆典仪式的地方。“那地方还保留有一些最原始血统的族民。”主演之一,具有毛利血统的克里夫&#8226;克蒂斯解释道:“唤醒对那片土地的神往是极其重要的。每一个参与影片拍摄的人都对这种膜拜情愫产生了共鸣,尤其我们的故事又与之息息相关。”
为了搭建亚高族村庄,工作人员分析了他们的生活方式以及他们当时赖以生存的那片土地。“亚高族手中的可用材料有限,” 艾默里克说:“他们只有猛犸象的骨头,尖牙和皮毛可以用来搭建他们居住用的帐篷。因为我们把他们想象成靠精神信仰赖以生存的人群,我觉得他们的住处应该是独特并且能够体现出他们的智慧和创造力的。”
艺术指导让-文森特&#8226;普佐斯把亚高族的住处设计成用骨头和皮毛搭制的,极具视觉冲击力和逼真的效果。“老妈妈居住的帐篷内部结构是用一万根猛犸象骨头搭建的,加上顶梁柱就是一副完整的猛犸象骨架。”他描述道:“我们用这个场景作为影片的开始,老妈妈在棚屋内主持着一场仪式。我们希望用它订下主基调。”
经过广泛的调查,从大量的考古学书籍中获得参考数据,普佐斯挑出了20付不同的猛犸象骨架。“我们仿制的骨头在比例上稍做了修改,这样在大屏幕上的视觉感受会更有冲击力。” 普佐斯说。“我们选择利用雕满部落符号的骨头和猛犸象头骨来装饰老妈妈的帐篷,加强精神信仰的神秘感,借此为影片的开始造势。”
用木头代替骨头的样子,普佐斯团队里的雕工们花了一个月的时间在南非开普敦制作基地制做骨架,与此同时,另一支队伍专心制作猛犸象的皮毛。完成后海运至新西兰的瓦纳卡镇,再由工作人员花费五周的时间组装。
维摩回忆当时的情况说:“我们有一屋子的人忙着打磨用木头做成的猛犸象骨架,都是在开普敦加工完成,零散海运到新西兰。我们还花了不少时间让海关相信,它们不是真的动物骨头。” 维摩笑道:“运输方面麻烦还不小呢,但至少保证了特别理想的结果。”
至于村庄内其余部分的装饰,工作人员在新西兰因地制宜地利用了很多不同的自然材料。“农场主们帮我们收集骨头,”现场装饰员艾米里亚&#8226;薇温德说:“我们还找到很多漂亮的海藻来装饰`帝克`居住的帐篷。
最关键的道具是普佐斯设计的白矛枪,将由帝克作为传位的象征物移交给他的继任。白矛枪必须设计得实用但又外观精致。最后的成品是一根大约六英尺长,顶端的象牙上刻满经文,拔掉象牙装饰可以变成利器的长矛枪。
和艺术设计相似,影片的服装设计欧达雷&#8226;迪克斯-米若和芮妮&#8226;阿普丽也极力希望演员的装束做到简单合理。迪克斯-米若从不列颠博物馆和开普敦的档案资料馆开始了她的研究。但是,她认识到,“在不列颠博物馆几乎没有什么可参考用于服装设计的资料。唯一的关于那个年代的直观记录就是一些在南非的画在岩石上的画。所以我们只能从剧本里汲取灵感并决定用不同的颜色来标识不同的部落:亚高族部落的颜色很简单,他们的颜色与他们生活的地形吻合。我们想到了一个办法,用非洲跳羚的皮毛保证了我们质料的来源。”
服装设计把亚高族的装束设计成抗寒,抵御环境侵害的样子。“没让他们穿鞋,” 阿普丽说,“他们一定很擅长利用动物的皮毛保暖,所以我们用厚重的羚羊皮代替猛犸象的毛皮制成他们的服装。由于天气的恶劣条件,抗寒装备已经比以前进步了很多。“我们也给演员们准备了能储备热量的装束,因为拍摄现场实在太冷了。” 阿普丽笑道。
戏服,发型,化妆和拍摄地点对史崔特上戏前的准备工作来说都不算难。“在新西兰的山顶上拍摄,由于你的发髻长到胸口使得你看上去很象一个猛犸象猎手,”他说:“我脸上自然长毛,再加上带顶假发,然后又把我的皮肤弄黑,感觉象常年在外被风吹日晒。尽管在每天早上上戏前得花点功夫,但看到结果觉得还挺值得的。”
至于袭奴军,迪克斯-米若设计的服装相对亚高族来说看上去更怪异但效果非凡。“我们运用了和亚高族使用的棕褐色完全不一样的色彩,”她说:“我们在麻布和毛织品中掺上了很多蓝色和红色,为了强调他们是生活在马背上的部落,我们用马尾为他们的装束增加效果。根据非洲部落资料,我们还为他们设计了面具和其他用麂皮制成的装束。
迪克斯-米若把汲自现存的非洲部落的灵感继续用来为亚高族勇士们旅途中遇到的其它部落设计服装。比如纳古族,郝大族和大河族。 “纳古族的色彩比较艳丽,我们还用土陶珠子制成的项链来代表这是个比亚高族更进步的部落。”她说。
最后一场戏,当达雷直面神灵和他的法师们,阿普丽设计了葡萄酒色的充满多种文化元素的装束,包括来自西藏和埃及的灵感。那些复杂的珠宝和化妆师托马斯&#8226;奈伦设计的脸部刺花,完善了整个角色的形象。
戏服往往能帮助演员进入角色,贴近并深刻理解角色的存在意义。“一穿上这些衣服你就觉得你已经进入了角色的世界,” 卡米拉&#8226;贝尔体会道:“它们会带你进入角色,你会发现自己的行为举止都跟原来的自己完全不一样了。”
除了主要演员的服装,服装设计组要为影片最后一场戏中近800多个群众演员负责奴隶的装束。先不说这数量,“阿普丽说:“这些服装我们无法批量订货,也无法用机器制作,全部装束必须手工完成,否则,会露馅。 我们几乎拥有一支专门在工作室做土陶,玻璃珠子的军队,并把它们一一缝制到服装上去,还有一部分的工人负责纺织品的制作和头饰。
六个部落的装束从头到脚都各具风格,服装设计组根据群众演员各自的尺寸,订做了近1000双戏鞋。
“我们还必须确保所有的服装看上去不能过于簇新,通过各种手段让它们看上去象穿过多年的旧衣服。阿普丽回想说:“这个电影的意境充满了野心勃勃的架势,外景地的选址令电影的拍摄工作更为艰难。但在南非和纳米比亚的工作团队真的都很棒!我们和当地很多技术熟练的工匠一起工作,比如鞋匠和帽工,他们的工作做得很到位。”
v摄制组从新西兰移师南非开普敦,这个国家已经为全球的电影制作行业提供了很多便利的基础条件。开普敦每年几乎和美国洛杉矶一样,要接待络绎不绝,来自世界各地的电影摄制组。剧组在那里找到了一片麦场和一间叫Table Mountain的摄影棚,适合用以拍摄达雷和伙伴们抵御食人鸟侵袭那场戏要用到的失落的山谷所需的内部结构。
Table Mountain的摄影棚和麦场位于开普敦的外围,为“失落的山谷”的构建创造了条件。为了制造高耸草从,使影片里食人鸟在攻击人类之前的埋伏藏身之处效果逼真,开普敦场地主管凯蒂&#8226;万弗和她的团队花了将近三个月的时间在麦场植草植树,养植草从。在开普敦,摄制组还在雷电城利用一间巨大的飞机修理棚完成了用陷坑诱捕剑齿虎的情节。
影片拍摄的最后一部份是在纳米比亚西南部的无人区沙漠完成的,包括质朴却闻名遐迩的Spitzkoppe岩石,艾默里克特别提到:“真是棒极了!只有纳米比亚你才会看到那样的风景和沙丘。”导演把丛山峻岭和沙漠的景观用合成镜头连了起来。 但是,每到一处,他总是被当地令人窒息的大自然景观迷得神魂颠倒。
让艾默里克对Spitzkoppe心驰神往的另一个原因,是因为他觉得Spitzkoppe岩石在他心里的地位之高是由于当年斯丹利&#8226;库布里克上世纪60年代就是用这块岩石作背景板摄制完成了电影史上具有里程碑意义的著名影片《2001太空漫游》。“这是个魔幻般的地方,不可思议!”
制作人维摩补充道, “Spitzkoppe 最与众不同的地方就是它真的会有共振产生,这种非同寻常的能量全世界只能在极个别特定的地方才能找到。无法统计它的数量,但真的让你感受到石体内这种能量的存在,很神奇!”
摄制组的人员被获准运用这块石头的神奇结构,在亚高族和纳古族相遇以及达雷开始掌握自己即将揭晓的命运的场景里都用到了它。纳古族是个发展完善的草原部落,以放牧和耕作为生。艺术总监罗宾&#8226;奥德说:“他们的村庄由建在岩石壁架上的小房子组成。小房子或者是建立在四面有框的架子上或者是建立在一个圆框上面,屋顶是用土砖盖住的。整个建造过程相当复杂。
Spitzkoppe是一处于公共境界的国家级历史文物,所有的地点征用费都上缴给当地社团。130名当地人被前期制作组聘用建造公路,用以圈围起影片中需要用到的跳羚和斑马的的栅栏。电影拍摄结束后,这些动物被捐赠给了当地社区正在筹备中的自然公园。
在靠近Spitzkoppe周围的地区没有酒店,整个摄制组驻扎在一个特别搭建的帐篷区,并有热水,电视和互联网设施。拍摄期间,每天6万公升的淡水从70公里以外的地方的运送进帐篷区。
影片中最神秘的情节就是达雷在沙漠中发现的失落的文明的见证。编剧科洛瑟之一说道:“当我们的英雄来到一个沙丘的顶端,他们看见了巨大的文明象征- 这些‘神峰’,这些神话般的金字塔,对他们来说根本无法相信出现在他们眼前的东西。他们长途跋涉的一部分原因也是渴望了解是什么样的文化让那么多人俯首为奴,要怎么样才能挑战一个这样的帝国。”
金字塔的建造位置在沙漠的七号沙丘处,靠近斯瓦科普蒙德(纳米比亚最著名的海滨城市)。艺术设计小组在这里建造了一个采石场,一个巨大的斜坡和法老的宫殿。 在利用直升机拍摄了人物长途跋涉的航拍镜头后,艾默里克想用相同的手段来展现金字塔的壮观场面。于是他让他的工作人员创建了一个巨大的金字塔模型,用spydercam摄影装置和一个遥控的有线摄象机拍摄。
工作人员在慕尼黑以1:24 的比例搭建起了金字塔的复制小样,宫殿,奴隶广场和靠近金字塔的尼罗河,再分15个海上集装箱运送到纳米比亚,总占地面积在100平方米左右。 spydercam摄影装置为导演自由地穿梭于小型复制现场提供了很大的便利,并提供了360度全方位的空中镜头,完成了影片的航拍场景。
“Spydercam摄影装置象直升机一样,”艾默里克说,它可以通过程序控制进行实时拍摄。灯光强度符合场景需要,背景部分又有真实的沙丘。对这组镜头的拍摄我相当自豪,因为它们是老式模型和现代技术的完美融合。
在与艾默里克合作了五次以后,电影摄影师犹利&#8226;斯特格特别珍惜与这位野心勃勃的导演在一个如此刺激的项目上一起工作的机会。“他是个特别合作的导演,有一流的想象力,” 斯特格说:“他在你还未加入他的项目前就已经把大部分他想要的东西搞定了,但又总是愿意倾听任何人给他的建议并做适当的调整。”
为了让影片保持其自然主义色彩,斯特格把摄像机和灯光的掌握控制在最小的范围内。尽量让自然光线为电影情节增效,带来最理想的感觉。“我们经常同时使用几部摄像机,这样当阳光充沛的时候,我们可以最大限度地利用它。”他说:“你的工作进度必须很快。我们通常会花数小时演习然后用三到四个摄像机进行拍摄,希望一次就顾及到所有的角度。
最后的元素就是《史前一万年》的配乐了。哈罗德&#8226;科洛瑟在此片的幕后担任的第三个角色是和他的同伴,作曲家托马斯&#8226;万德共同谱曲完成的 。 两位作曲家和导演密切合作,希望从音乐的角度捕捉独特的故事情节中行为和感情上的表现。
导演说:“故事是个精彩的英雄神话,音乐必须和它相呼应。但它又包含很多部落民族的元素,大喇叭,和声和鼓类乐器。拍电影还有个很有乐趣的地方就是期待音乐怎么和你的人物形象相吻合。奇妙的时刻就是当你把第一部分用交响乐表达出来以后,你就觉得万事顺意。
对艾默里克来说,最后的剪辑合成是把每一个辛苦创造的碎块拼凑在一起,这也是享受所有艰苦劳动后的最佳回报。“拍电影是件错综复杂但又特别好玩的事情。完成一个电影之前要顾及到方方面面,我特别喜欢让自己身在其中并乐此不疲。我也总是喜欢在整个过程中发现或发明更有意思的做法。不过不管科技有多发达,最关键的还是故事里的角色。再精彩纷呈的情节如果没有人物的穿针引线,也就索然无味了。”
通过一个年轻人一段非凡的旅途,《史前一万年》的故事中延伸开了好几段不同意义的主题,包括英雄主义本身的定义、人类团结的力量,以及人与人之间存在的阶级关系。“每个人都得自己决定自己想生活在一个多大的人际圈里,” 艾默里克说:“是仅仅自己爱的人,他的家庭还是一个更广阔的范围?我们故事里的英雄必须通过一段千辛万苦的长途跋涉才能找到答案。他必须从一个懵懂自私的孩子迅速成长为一个领袖。关键就在于他如何圈定自己……你在你的圈子里包容了多少人。”
发布者:Mtime (2008-02-29 09:53:06)
划时代的一次旅程
寻找失落的世界
罗兰德&#8226;艾默里克在自己的职业领域里一直试图挑战视觉效果的所有可能性。在他之前的作品中,类似《独立日》中白宫爆炸等惊心动魄的大场面以及《后天》中滔天巨浪淹没纽约城,已经有目共睹。艾默里克一次次地释放着他无穷的想象力并借助不断发展的技术让他在《史前一万年》里的创作再一次过足了瘾。
艾默里克力邀曾经和他在《哥斯拉》和《后天》中合作过的视觉效果指导卡伦&#8226;古丽卡丝与他第三度联手,监督本片大场面的制作。“卡伦是我合作过的最有独创精神,并最擅长开发视觉效果的专家,”导演说,“对她来讲,没什么是不能做到的。我深信,有了她的协助,我最具野心的想象也能在银幕上得以实现,甚至比我当时的想象更壮观可信。”
本片最繁重的工作就是对远古时代各种动物的再现,巨型猛犸象,剑齿虎和食人鸟。 艾默里克要求这些动物有最逼真的动作,接近已然进化至今的它们的后代。“我们参考了大量的大象,老虎和鸵鸟的影像资料,”他说,“主要的问题在于人们没有见过真实的猛犸象是什么样的。它们是一种极其独特的动物。只能从现有的影像资料里汲取可利用的东西。”
而最具挑战性的是近距离再现远古动物的毛发,猛犸象的毛发长而纠结,食人鸟的羽毛和剑齿虎遇水后的处理。 “我们基本上必须重新创造新的毛发形态的模块,才能使这些动物在影像里想真的一样,” 艾默里克说:“要做好还真不容易,我们在英国聘用了两个专业公司完成这部分的工作,现在你能看到的动物真实得触手可及。
两年前,主体摄影还未开始,古丽卡丝就已着手开始影片视觉效果方面的工作,将剧本根据场景的需要做分解修改。把故事中想象的动物和场景变成具体可见的雏形,设计初步的草图(扫描雕塑品到电脑里)和制作模型。她的工作重点就是本片最出彩的三个环节:捕杀猛犸象,食人鸟的追逐场面,达雷和剑齿虎对峙的场景。
古丽卡丝创建了一个图库,收集了各类照片和从电视里截取的CG影像,作为本片史前动物创造的资料源泉。她还特地走访了位于美国洛杉矶市汉考克公园内的拉布雷亚沥青坑,在那里寻找到了对研究猛犸象极有参考价值的资料。另外,古丽卡丝还去了南非德班的野生动物园,拍摄了很多野生动物的高清影像资料,例如非洲狮,老虎,美洲豹,大象和鸵鸟。她的这些资料给动画组的工作人员提供了以各种角度研究动物的可能性。
古丽卡丝最大的工作挑战就是食人鸟的制作:不会飞的食肉动物,长有巨嘴。这种动物的创造灵感来自于南美现存的一种动物。“它们身形庞大,” 古丽卡丝说,“我们已知的是一只鸵鸟的奔跑速度以及它的一只脚所能带来的杀伤力。于是我们把已掌握的确凿事实和可参考资料相结合,比如说食人鸟和恐龙一定有某种关联,这样一来,就可以越来越准确地把已经混合的动物形象特征经过想象勾勒出来。”
将动物的动作完美化需要跟导演艾默里克一起,通过对它们的设计进行千锤百炼才能完成。“这是个彻底的挖掘过程,” 古丽卡丝说,“你不断地修改再修改,直到完全符合为止。这部电影的制作建立在无数的创新、通力合作以及无尽的开拓上。罗兰德给我支了很多招,同时又没有给我任何约束。”
一旦动物的主体形象设计确定,古丽卡丝手下的18人团队,包括动画师,道具、布景制造等工作人员便开始了前期的造型视觉化工作,为所有的情节制作3D效果的串联板。“比如说,达雷和剑齿虎在陷阱里的那场戏,我们制作了整个陷阱的3D模型和演员的模拟形象,并从鸟瞰的角度设计了老虎纵身一跃的动作场景, 以此确定所有该有的动作。” 古丽卡丝解释道:“然后我们设计调整各个镜头的角度,通过视觉效果编辑史蒂文&#8226;庞和视效小组的努力,展示出预演的效果。我们对每个镜头的分割进行逐一地讨论,对每个演员实地拍摄时需要做到的具体内容进行确定。”
前期视觉效果制作渐渐成为实地拍摄时演职人员最为依赖的宝贵工具。“我总是让演员在场景搭设前先观看视觉效果演示,这样他们就能提前知道即将在他们身边发生的各类情况。”艾默里克说。
对摄影指导犹利&#8226;斯特格来讲,视觉效果演示为灯光的设计也起到了非同一般的作用。 “视觉效果演示对一个特定场景的拍摄有实际指导作用,” 斯特格肯定地说:“当然,最后拍摄完成的效果正因为有了视觉效果演示的指导,才让我们在真实拍摄的过程中做到扬长避短,结果自然就更为出色。”
艾默里克的民主工作作风使得古丽卡丝的创作团队在想象的王国里自在徜徉,导演和视觉效果工作组保持着轻松密切的工作关系。“我们经常讨论他们的新想法并设法把这些想法运用到工作中去。”她说:“这使得我们的工作质量更上一层楼。视觉效果艺术家们的使命责任感令他们经常觉得自己也有讲故事的责任。”
在拍摄现场,古丽卡丝和她的团队参与到演员和其他工作人员们中,用工具丈量计算虚拟的史前动物角色正确的方位和运动轨迹,并用蓝色标记出需要后期用数码制作的影像替代的部分。“ 食人鸟的动作场景里,我们把一个涂成蓝色的食人鸟头固定在标尺的一头,按照既定的轨迹运动,方便摄影机取景拍摄。”她解释道:“而在拍剑齿虎的动作场景时,我们按1:1的尺寸把老虎画在标记上,以此为依据取景。如果镜头出现偏差,会给后期制作带来很大困难。有了模拟的动物配合,演员的表演就有了互动,导演也能拍摄到他想要的画面。”
对片中年轻的演员来说,与视觉效果专家们的合作是段极其有趣的经历。“这是个好好利用你的想象力的绝佳机会”,史蒂文&#8226;史崔特说:“它给了你足够的空间,任你发挥而不会受任何阻碍。在拍摄捕杀猛犸象的那些场景的时候,享受自由发挥,与不存在的东西做互动是件刺激的事情。”
我想,你说的刃齿虎的"最大的化石",就是科普论文里的截图吧。
就是这个。44-45厘米的大头骨。科普论文里有贴南美洲的刃齿虎全身骨骼图,你可以按照它的头骨是45厘米来算算肩高多大。我算的结果就是1。2米或者1。3米的样子。你如果有更大的化石能证明出来它的肩高有1。6米,欢迎提供。谢谢。
关于体重,很多都是推测。按照雄东北虎的模型来推测的话,最大的刃齿虎大约470KG。
(本文为化石网论坛精华帖,由网友绫崎飒提供,仅供大家学习参考。编辑:常君滢)
2022-11-30
-

真正的世界末日
有鳞目(Squamata)是现今爬虫纲中最大的目,种类能多达9千种之多,可以肯定的是处于极盛的黄金时期。有鳞目(Squamata)被认为在中生代进行辐射性演化,在侏罗纪已出现了冠层的种类,蛇类及蜥类在白垩纪时期分化有鳞目(Squamata)是现今爬虫纲中最大的目,种类能多达9千种之多,可以肯定的是处于极盛的黄金时期。有鳞目(Squamata)被认为在中生代进行辐射性演化,在侏罗纪已出现了冠层的种类,蛇类及蜥类在白垩纪时期分化。普遍认为大批有鳞目种类能避过晚白垩纪至古新世之间的大灭绝(K-Pg),除了那些海上称王的沧龙总科以外。但是近来北美发现的化石显示,有鳞目在这次大灭绝也几乎全军覆没,绝种的百分比竟多达83%,以下简略介绍一下北美的有鳞目化石。
Polyglyphanodontia是一类已灭绝的蜥类,牠们的下颌联合呈V形及具较长的夹骨,牙齿呈三齿尖,牠们现被认为是不属于硬舌亚目(Scleroglossa)。已知的种类包括Polyglyphanodon sternbergi,Chamops segnis,Leptochamops denticulatus,Meniscognathus altmani,Haptosphenus placodon,Stypodontosaurus melletes,Peneteius aquilonius及Tripennaculus。此外以下的蜥类也是属于Polyglyphanodontia:
Socognathus brachyodon有粗状的下颌及厚重的联合,附齿尖缩小,Socognathus brachyodon有紧密排列的球状牙齿。
纤细奥巴蜥(Obamadon gracilis)是小型蜥类,有较高而纤细的牙齿,这属名亳无疑问是纪念当今美国总统。此外在科罗拉多州及萨斯喀彻温省也发现了Polyglyphanodontia的化石。
有三种石龙子下目(Scincomorpha)化石被发现,Contogenys sloani过去被认为是属于夜蜥蜴科(Xantusiidae)茎层种类,但是现在被划归石龙子下目的球蜥科(Globauridae)。
Lonchisaurus trichurus有直长的齿骨,有数目众多且排列紧密的牙齿,齿尖较钝,Lonchisaurus trichurus在石龙子下目的位置不明。
谷氏爱斯特蜥(Estescincosaurus cooki)在石龙子下目的位置也是不明的,三种石龙子下目中只有Contogenys活到古新世。
Exostinus lancensis是北美发现的异蜥科(Xenosauridae)茎层种类,Exostinus lancensis在古新世时期以后再没有化石记录,但是该属另一种类Exostinus serratus则被发现于科罗拉多州,化石属早渐新世时期。
已绝种的蛇蜥科(Anguidae)种类包括奥达克蜥(Odaxosaurus piger),牠是原始的雕蜥亚科(glyptosaurine)。Litakis gilmorei及Colpodontosaurus cracens在蛇蜥类(Anguimorph)的亲缘关系不明,此外还有侧褶蜥属未定种(Gerrhonotus spp)化石被发现。
当地发现了四种巨蜥类包括博格副毒蜥(Paraderma bogerti),怀俄明副萨尼瓦蜥(Parasaniwa wyomingensis),古萨尼瓦蜥未定种(Palaeosaniwa sp)及巨大塞姆蜥(Cemeterius monstrosus),牠们都属于巨蜥类的茎层种类。
怀俄明副萨尼瓦蜥(Parasaniwa wyomingensis)
塞姆蜥属(Cemeterius)有短而厚重的颌骨,牙齿粗状,牙齿舌面膨涨且没有条纹,塞姆蜥属(Cemeterius)是大型的巨蜥类。
古萨尼瓦蜥(Palaeosaniwa)也是白垩纪的大型巨蜥类, 当地发现的古萨尼瓦蜥未定种(Palaeosaniwa sp)估算吻部至肛门的总长为82厘米,体重可达6公斤。
加拿大古萨尼瓦蜥(Palaeosaniwa canadensis)
兰斯副鬣蜥(Pariguana lancensis)是北美已知最古老的鬣蜥种类,从化石来看副鬣蜥属(Pariguana)比亚洲的原始鬣蜥要高等,副鬣蜥属(Pariguana)的化石发现显示鬣蜥类早于白垩纪时期已从亚洲扩散至北美。
残酷拉米亚蜥(Lamiasaurus ferox)有高而弯曲的齿冠,齿尖下有颈缩,牙齿有舌脊,残酷拉米亚蜥(Lamiasaurus ferox)保留了短小的夹骨,因此拉米亚蜥属(Lamiasaurus)被认为是鬣蜥类的茎层种类。
在北美发现了三种蛇类包括原始蛇类Coniophis precedens及两种真蛇下目(Alethinophidia),包括粗状刻耳柏洛斯蛇(Cerberophis robustus)及另一种未定种名的小型品种。Coniophis保留了蜥类的头部及蛇类的躯体,牠可能还保留了后肢残余。
Coniophis precedens
粗状刻耳柏洛斯蛇(Cerberophis robustus)有宽阔的脊椎,脊椎骨宽达17 mm显示体长超过1.7米,重量超过2.9公斤。刻耳柏洛斯蛇属(Cerberophis)在当时而言已算是大型蛇类,牠的体型足以吞食当时的哺乳类,甚至是刚孵化出生的恐龙。粗状刻耳柏洛斯蛇(Cerberophis robustus)比早古新世最大的陆上蛇类Helagras prisciformis大得多(Helagras prisciformis全长超过950 mm,重量超过520 g),刻耳柏洛斯蛇属(Cerberophis)最初还被认为是归属蟒科(Boidae),
Helagras prisciformis
在北美发现了多达27种类蜥类及3种蛇类化石(有许多尚未被定种),化石全属晚麦斯特里希特阶(Maastrichtian),竟只有Exostinus,侧褶蜥属(Gerrhonotus),奥达克蜥属(Odaxosaurus),Contogenys及原始的蛇类Coniophis支持至古新世时期,根据化石记录来看多达83%品种是绝种的,可见大灾难对当时有鳞目带来毁灭性的打击。Polyglyphanodontia完全灭绝,至于巨蜥科(Varanidae),美洲鬣蜥科(Iguanidae),石龙子科(Scincidae)及壁虎下目(Gekkota)等的茎层种类也全军覆没。不过与此同时亚洲的种群却向北美及其他地域扩散,欧洲发现了古新世的正蜥科(Lacertidae)化石,南美及印度也发现了古新世至早始新世的真蛇下目化石等都显示有鳞目在大灾难以后死灰复燃,在古新世时期进行辐射型演化。
(本文为化石网论坛精华帖,由网友Sargon提供,仅供大家学习参考。编辑:常君滢)
2022-11-30
-
江苏句容鱼的目科分类
江苏句容早三叠纪出产鱼结合,凡是在化石网上溜达过的都知道,但是其中并不是一种科目的鱼类存在,其实这个问题大家也都知道,但是小弟愚钝,竟然在论坛上混了好几年今天才将将能把句容结合鱼大致分个科目来,再江苏句容早三叠纪出产鱼结合,凡是在化石网上溜达过的都知道,但是其中并不是一种科目的鱼类存在,其实这个问题大家也都知道,但是小弟愚钝,竟然在论坛上混了好几年今天才将将能把句容结合鱼大致分个科目来,再次感谢一直得瑟我并帮助我学习成长的良师损友“无所谓”童鞋。
东昌青山早三叠纪下青龙组上部发现的鱼类 ,共计有2目,2科, 6属, 8种。至此,本人将将能够鉴定到科,至于属和种,感兴趣的童鞋继续深造吧!
软骨硬鳞鱼次纲Chondrostei
裂齿鱼目Perleidifo rmes
裂齿鱼科Perleididae
总结最简单的鉴别方法是sop下鳃盖骨>pop前鳃盖骨>op鳃盖骨,这里面op最小;口裂很深,发现没有?
当然还有就是看上去的感觉,裂齿鱼的脑袋是方的!!
新鳞鱼次纲Neoptergii
半椎鱼目Semio no tiformes
半椎鱼科Semionot idae
半椎鱼科的鳃盖系统是op>sop>pop.还有就是口裂小,且有sbo次眶骨,这个是半椎目的主要特点,其他还有许多不同,大家自己对比吧!
这种鱼类的头部看上比较瘦长,且嘴部尖锐,下颚部有块Ang-隅骨(白话就是下颚骨分成了两端,牙齿着前半段)
(本文为化石网论坛精华帖,由网友蛰无敌提供,仅供大家学习参考。编辑:常君滢)
2022-11-30
-
共享的三叠纪鱼类(云贵地区)资料
古鳕目在地史中存在时间很长,从泥盆纪到白垩纪都有,而且在全世界范围内都有古鳕目的分布。在云贵三叠纪地层中也广泛分布着古鳕目的鱼类。少鳞贵州鳕口裂长、上下颌为典型古鳕型,鱼体比较大,长纺锤形。化石图古鳕目在地史中存在时间很长,从泥盆纪到白垩纪都有,而且在全世界范围内都有古鳕目的分布。在云贵三叠纪地层中也广泛分布着古鳕目的鱼类。
少鳞贵州鳕
口裂长、上下颌为典型古鳕型,鱼体比较大,长纺锤形。
化石图片:
兴义鱼
弓鳍鱼目
弓鳍鱼亚目
真颌鱼科
兴义鱼科
鱼个体较小。身体长纺锤形。头骨表面有很薄的硬鳞层。上、下颌骨细长, 后部微增高。口裂长, 口缘有锥状牙。下颌的冠状骨发育且具有几排粗钝的牙。辅上颌骨小而细长。鳃盖骨比下鳃盖骨大。
(图片来源:化石网)
(图片来源:Zero)
真颌鱼
鱼中等大小, 纺锤形。头短而高, 吻钝圆。头骨表面有薄的硬鳞层和瘤状饰纹。顶骨小而方形, 额骨大而长。鼻骨小而长方形。
贵州真颌鱼
优美贵州弓鳍鱼
弓鳍鱼科
贵州弓鳍鱼属
早期弓鳍鱼
肋鳞鱼
东方肋鳞鱼,大家估计不陌生,鳞片特征比较明显。描述略。。。。
空棘鱼
空棘鱼目,大家熟悉的矛尾鱼
短体鱼
卢加诺鱼亚目
卢加诺鱼科
鱼很小, 身体短而高。鳃盖骨和下鳃盖骨三角形, 前者比后者大。背鳍颇大而三角形, 位置很靠前, 起点与腹鳍起点相对。臀鳍较大而非常后位, 起点对着背鳍的终点。尾弱半歪型, 尾鳍正型, 有很弱的尾裂。所有的鳍棘鳞显著。鳍条近端不分节, 远端分节并分叉。硬鳞较厚而表面光滑。身体前侧面有两列狭而高的纵列鳞。
扁体鱼
扁体鱼亚目
结构特征与古鳕很相似,就是体态有所区别。在浙江地区有扁体鱼的发现。但贵州的资料很少,我也暂时没有找到更多关于扁体鱼的资料。但我个人对国外扁体鱼标本比较和对贵州见到的“弓背鱼”和“团鱼”两种鱼类化石标本来看,此两种应该可归入扁体鱼亚目(个人观点,应该是首次提出吧,已开始搜集、整理和编写相关资料了)。
扁体鱼复原图(用Adroichthys)
比耶鱼
软骨硬鳞鱼超目
比耶鱼目
比耶鱼科
比耶鱼属
龙鱼
以前介绍过,辐鳍鱼亚纲 ——软骨硬鳞目——龙鱼目。龙鱼身体延长,吻极其突出,身体一般有4行骨质硬鳞(如甲片,背部、腹部以及身体两侧各一行),其他部位无鳞或薄鳞。尾鳍对称,背鳍在后臀鳍上方。
亚洲鳞齿鱼
体型为高纺锤形,头骨外部骨片和鳃盖的表面具有突起。鳃盖骨略呈长方形, 上端稍窄, 其位置略向前倾。鳞片较厚大, 具有厚的阴光盾, 表面光滑, 后椽具梳状齿, 躯干前部之侧麟较大,长方形。
鳃盖骨:
与真颌鱼头骨的对比
特别插入云南罗平鱼类介绍,罗平生物群中发现大量中三叠纪的鱼类化石,意义重大。简单上几种鱼类
全骨鱼类
裂齿鱼
肋鳞鱼
真颌鱼
龙鱼
不是秀丽兴义鱼
Saurichthys yunnanensis sp.nov
Luopingichthys bergi gen.n.sp
Gymnoichthys inopinatus gen.n.sp.n
弓背:Luoxiongichthys
(本文为化石网论坛精华帖,由网友流水无心提供,仅供大家学习参考。编辑:常君滢)
2022-11-30
-
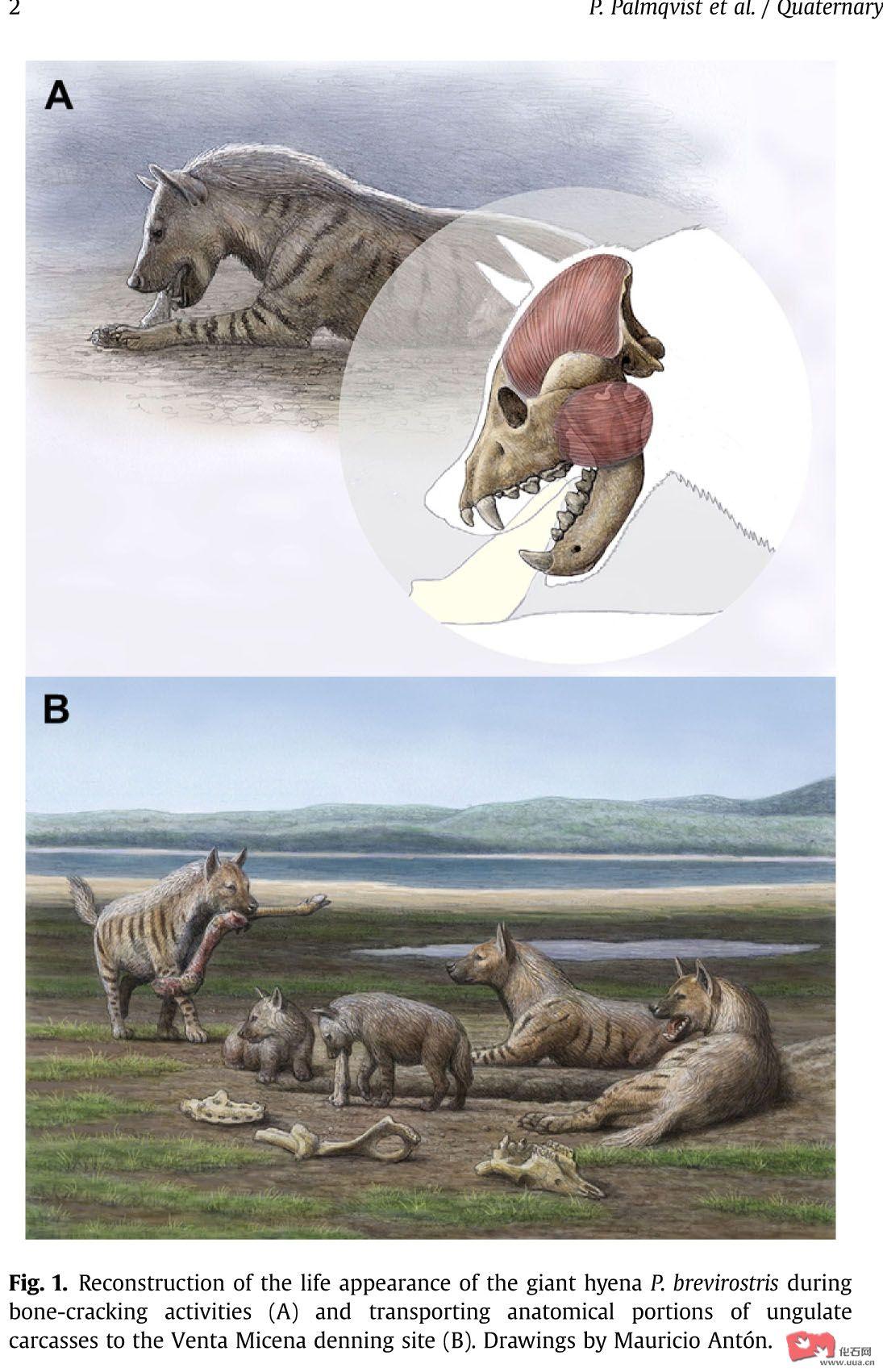
怪兽复活之冰河春秋——浅谈早更新世大型肉食动物
介绍:硕壮鬣狗,欧美洲虎,欧洲的巨颏虎属,锯齿虎属桑氏硕鬣狗(PACHYCROCUTA BREVIROSTRIS),在欧洲,亚洲(中国周口店)和非洲东南部都有分布,是最大的鬣狗科之一,中国发现的巨恐鬣狗是中鬣狗科的。这个动物的
介绍:硕壮鬣狗,欧美洲虎,欧洲的巨颏虎属,锯齿虎属
桑氏硕鬣狗(PACHYCROCUTA BREVIROSTRIS),在欧洲,亚洲(中国周口店)和非洲东南部都有分布,是最大的鬣狗科之一,中国发现的巨恐鬣狗是中鬣狗科的。
这个动物的大体描述英雄王和小超已经都说了,这里补充一些更详细的数据
桑氏硕鬣狗的正模是在法国发现的,但是偏部欧洲了
头骨正模就有基底长322mm,这个已经是雄非洲狮的范围内了,周口店也有一个颅全长351mm,基底长315名木的标本,此外322mm的估测全长357mm,而且正模还并不是特别大的,至少周口店和欧洲其他地区都有很多下颌骨接近这个水平,超过的也有,而西班牙Incarcal还发现过肱骨长度可达310mm的桑氏硕鬣狗,基本上肱骨长度可以匹敌当地发现的阔齿锯齿虎(论文里面说那个硕鬣狗比阔齿锯齿虎长,其实不对的,Incarcal的锯齿虎肱骨有320mm,但是基本上差不多了)
再补充一些
按照颅基长度,桑氏硕鬣狗体重可以达到190kg左右
但是Turner提出桑氏硕鬣狗的肱骨和股骨到不了190kg的鬣狗的水平,而桡骨,胫骨都相对来说还要更短,因此桑氏硕鬣狗肩高估测在1米左右
以下是桑氏硕鬣狗和一头肩高85厘米的大个体的斑鬣狗的比例
稍微说一下印度的桑氏硕鬣狗,巴基斯坦Haro河发现的桑氏硕鬣狗(文中别名叫短面巨鬣狗)的洞穴,包括一些有幼年个体,确认的特点是巨大而且很高的下颌骨,还有M1下原尖呈现球状,下原尖和下前尖相对短但是冠尖突出,P2和P4相对M1较长,硕鬣狗演化渠道是通过印度(siwalik)达到中国,硕鬣狗在巴基斯坦有啃老鼠的,有啃郊狼大小的犬类,鼬类(狼獾大小),不明猫科动物,獴
啃的素食动物有牛头骨,为了吃脑髓,颅基部分几乎被啃掉了,长骨也只剩下骨头末梢,不过有些特别大的骨头,长骨中段没有完全被啃掉,倒是末梢有些被损坏了
参考的现代鬣狗科的三个种的联合计算,这次使用更多的公式
上面估测的108kg是平均的成年偏大个体,最大的西班牙标本按它的肱骨(310mm)算大概155kg
肱骨和桡骨比例和洞穴斑鬣狗最接近,而胫骨和股骨,四肢综合的比例和综鬣狗最接近,牙齿也是比较接近现代鬣狗属或者洞穴斑鬣狗,距离现代斑鬣狗貌似差距最大
对要自己比较大小的人可以参考以下《撒哈拉南部动物志》里面记载的鬣狗科的肩高,体长对应的体重数据
桑氏硕鬣狗很可能起源于亚洲
300~50万年前:亚洲的Siwalik和周口店,爪哇,Transbaikalia,外蒙古等地都特有发现
300~150万年前:从亚洲进入非洲动物,扩张到非洲南部,在东非发现过很大的下颌骨(这里面不贴了,主要讲欧洲的动物)
160~50万年前:欧洲的主宰食腐动物以及优势的鬣狗科动物,在更新世早期进入欧洲以后取代了P型更新世鬣狗(Pilocrocuta Perrieri),但是在更新世晚期后者回归欧洲,之后由于锯齿虎属和巨颏虎属被狮子取代,靠食剑齿虎科无法利用的尸体剩余为生的桑氏硕鬣狗失去了重要的食物来源,由于其体型过大,行动能力不佳,和早期人属动物竞争加剧,肢体比例不适合长途跋涉等原因,退出历史舞台,P型更新世鬣狗回归欧洲,并且和洞斑鬣狗将桑氏硕鬣狗取尔代之
Homotherium latidens(阔齿剑齿虎),Acinonyx pardinensis(巨猎豹),Megantereon (锯齿虎属),Panthera "onca" Gombaszogensis(欧美洲虎)
Panthera leo (狮子,特指亚种古狮和洞狮),Xenocyon Lycanoides(欧洲豺犬,其实就是现代非洲三色豺的同属,早期更新世进入欧洲的亚洲三色豺犬演化的,有的时候也称Asian hunting dog),Xenocyon Falconeri(法氏豺犬,体型和现代的大亚种狼差不多,头骨有24厘米多的)
Megalceros(大角鹿属),Cervus(鹿属),Alces(驼鹿属),Dama (黇鹿属),Rangifer(驯鹿属)
桑氏硕鬣狗喜欢吃动物骨骼的长骨,也啃脊椎骨,颜色越浅的保留数量越少,越深的被啃的也越少
桑氏硕鬣狗的牙齿比起现代斑鬣狗外更适合啃骨头,而且咬力也相当的大,但是它肢体比例不适合长途跋涉寻找尸体,因此往往是跟着锯齿虎等的屁股后面捡剩肉,当然遇到巨颏虎这个大小的,估计就可以之际赶跑了,不过它单打独斗打不过大型的欧美洲虎,可能也面临和人类的竞争(在一些有利的情况下早期人类会猜测会用丢石头的方式驱赶掠食动物并夺取他们的食物),桑氏硕鬣狗也可能捕猎,例如攻击大型动物的行动迟缓的幼年个体,现代不同地区的斑鬣狗也有偏向食腐肉/捕猎的
桑氏硕鬣狗的咬力和颌骨,牙齿结构更加接近鬣狗属,而不是斑鬣狗属,但是咬力明显比他们都大的多,也更特化于啃咬骨头,估测臼齿咬力除了巨恐鬣狗和一些熊之外,几乎陆地肉齿目很少可以匹敌了。
PS:按照头骨模型(dry skull method)体型足够大的大猫的末端咬力不比鬣狗小,但是就是大猫一般不用那么大的力拿臼齿啃骨头而已。
以下都是在希腊马其顿发现的肉食动物:下图地图
其中早更新世这个阶段发现的就有Canis arnensis, Lycaon lycaonoides, Vulpes praeglacialis(狐狸),Pliocrocuta perrieri, Pachycrocuta brevirostris.
区别桑氏硕鬣狗是根据其P4上裂齿的大小明显超过斑鬣狗和更新世鬣狗属的范围,当地也发现的相当大的洞熊,斑鬣狗古代欧洲熊类和狮子,但是那些都是中晚期更新世的材料了
Pliocrocuta perrieri的头骨和下颌骨,和斑鬣狗的比较,上裂齿P4切割能力较差,因此更倾向于食腐肉,下裂齿M1有明显的下后尖,下裂牙远中部夸大,上裂齿里面P2是P4长度的一半左右,而在斑鬣狗种P2远远小于P4,在希腊发现的Pliocrocuta perrieri比起欧洲其他地方的标本显得比较原始,它下颌骨和斑鬣狗的对比图,其实末端的牙齿没有斑鬣狗那么发达
现在再来看看欧洲其他地区的动物分布
下图:其实西班牙崎地区给的表格化石更加全面(有些英文翻译前面已经给了不在重复)
除了马(Equus),三趾马属(Hipparion),貉属(Tapirus),猪(Suidae)牛等还包括了古老古菱齿象(Mammuthus antiquus),长毛象(Mammuthus primigenius),南方猛犸(Mammuthus meridionalis),嵌齿象科(Mammutidae),嵌齿象科(Gomphotheriidae),古河马(Hippopotamus antiquus),双角犀牛属(dicerorhinus),美洲野牛属(Bison),牛属(Bos),水牛属(bubalus),西方狍(capreolus), 真枝角鹿(Eucaldoceros), 原牛,古长颈鹿(Pliotragus),羊牛属(Ovibos) 还有很多古羚羊类(Gazella,gazellospira)和很多其他的素食动物,当时早更新世的动物和非洲很像呢,虽然中晚期就没有这些特点了
praeovibos(原牛类),Hemitragus alba(阿氏塔尔羊),大型的南方猛犸可以达到6000kg,古河马比现代的河马大很多,可以达到3000kg,Megantereon whitei(怀特巨颏虎),法氏三色豺犬(canis falconeri),意大利熊( ursus etruscus),意大利狼(canis etruscus),狐属(vulpes),猞猁属 (lynx)
下面是英国早期中更新世的动物分布,外加德国的
英国的Pake field
大型肉食动物锯齿虎和巨頦虎,狮子共存以外,也和巨猎豹,欧美洲虎,狼,欧洲三色豺犬(也称欧洲山狗),藏马熊,几种欧洲的古代棕熊洞穴斑鬣狗,上新世鬣狗属和桑氏硕鬣狗等共存过。同时其的草食动物也可以看一下表格,伊特鲁尼亚犀(Stephanorhinus), 草原猛犸(Mammuthus trogontherii),古老古菱齿象(Palaeoloxodon antiquus),现代河马(Hippopotamus amphibius),三种大角鹿属,两种马属,黇鹿,马鹿,北美野牛属(Bison)和野猪(Sus scrofa)
在德国的动物
三种马属,草原猛犸,古老古菱齿象,河马属,西方狍,马鹿,两种大角鹿属,野猪,伊特鲁尼亚犀,驼鹿和人属不确定种。
和英国的总类很相似,因为那个时候没有英吉利海峡,这些动物可以到达英国,直接走过去
Group1: 温度和现代相等
Group2:夏天比现代热,年代和1一样,包含很多大型动物
Group3/4:夏天比现代热
Groups 5/6/7:其购比较多变
英国的大角鹿和锯齿虎
注意:那个锯齿虎的测量尺其实是5厘米,图上10厘米是打错了,否则这个下颌骨长度近40厘米,头骨近60厘米,明显是一个1吨多的锯齿虎的头骨
此外Pakefield还有很大的狮子呢,那个下裂齿33mm,大概也就俄国Ural的洞狮和法国洞狮有那么大了。
英国的欧美洲虎肱骨末梢
锯齿虎属
狮子
当然也有人推测有很大巨型鬣狗科的动物
材料居然是一坨粪便化石,给出了和现代非洲斑鬣狗粪便的比例
当然还有测量表格,确实明显超过现代鬣狗(野生+圈养)的范围了,最下面的那些最大估测都是桑氏硕鬣狗的粪便。
欧洲在西班牙,早更新世晚期,也有桑氏硕鬣狗和熊类,豺狗,欧美洲虎一起,这里的桑氏硕鬣狗正处于更新世过渡期,也许它们是欧洲最晚期的种群之一了。
测量数据:PS 14521: P2/ L = 22.4, P2/ W = 15.2, P3/ L = 27.4, P3/ W = 20.1, P4/ L =43.4, P4/ Lp: 14.1, P4/ Lm 5 17.9, P4/ W 5 24.6; IPS 40627: I/3 L = 8.1, I/3 W = 11.1; IPS 18381: P/2 L = 19.4, P/2 W = 14.8; IPS 40625: P/4 L = 28.2, P/4 W = 16.6; IPS 13255: I2/ L =8.2, I2/ W = 9.45.
鉴定特征:上裂齿p4是唯一有中齿尖突出的牙齿,末端齿尖高度超过中齿尖,P2有很小的前齿尖,P3相对比较宽,P4相当的大,上原尖很发达,和P3的上原尖一样高,上前尖很也很高。
还有当时同地点的一些古熊类的化石
现在介绍一下锯齿虎属,锯齿虎属牙齿的特点,大管家已经详细的说了,我就不画蛇添足了,尤其是我对牙齿不很熟悉
锯齿虎图片里面西班牙的Incarcal阔齿锯齿虎的体形比大型亚种雄狮子稍小,肱骨320mm,但是阔齿锯齿虎是有一些较大的标本,例如1906年发现的阔齿锯齿虎的肱骨就有380mm长,末梢102mm,比同长度的巨型剑齿虎的肱骨更粗了10%左右,桡骨也基本上大概比毁灭刃齿虎更长些,圆锯锯齿虎似乎更大,头骨也更粗壮,在英国也发现了宽度可至103mm的肱骨末梢。
西班牙发现的阔齿锯齿虎:同长度下肱骨比狮子细,头部占身体比例比狮子小,爪子比狮子短,但是颈椎骨比狮子粗,手掌也狮子强壮
锯齿虎的骨骼有豹属,猎豹属和斑鬣狗的混合特征,其脊椎骨前段部分最类似美洲虎. 阔齿锯齿虎在前肢II-V指上有较小的爪子,在I号指上有较大的爪子,爪子介于猎豹属和豹属之间,推能有伸缩能力,前肢的手掌部分相当的粗壮,超出狮子。锯齿虎的腰部脊椎骨相对坚挺,可以有效的为前肢提供一个稳定的发力平台,也可以在帮组承受猎物挣扎时导致的压力。锯齿虎的符合地栖性大猫的特征,是用前肢的方式类似于刃齿虎(两者的前肢上发现类似的使用性损伤)锯齿虎的后肢脚部的关节比豹属的左右转动幅度更大,这样可以帮组锯齿虎以半直立姿式控制大型猎物时避免摔倒。锯齿虎的前肢肩膀部分高于后肢/骨盆部分,此处很类似斑鬣狗,锯齿虎的短距离加速能力较差,后肢比前肢弱,但是它在长途奔跑的时候能量利用效率高于豹属
阔齿锯齿虎的体格比传统的刃齿虎和现代狮虎纤细些,但是仍然能采用前肢压制猎物,安东和Turner推测锯齿虎联合狩猎,直接在当场放倒猎物。虽然北美的血刃锯齿虎确实有吃小猛犸象的记录,阔齿锯齿虎的主要猎物其实应该不是那些它们无法压制的成年大象和犀牛,而是马,牛或者鹿一类的。
安东在1999年描绘了锯齿虎如何使用锯齿的,安东描绘了锯齿虎的7块颈椎,除了第7号之外其余的都相对豹属的更长,颈椎有一些有类似刃齿虎的特征,锯齿虎的冠状突和角突的退化没有费氏巴博虎等巴博虎亚科和刃齿虎那样明显,其嘴巴能长开的角度也略小于刃齿虎,不过锯齿虎的锯齿远比刃齿虎要短。锯齿虎的脖子可以进行很大幅度的移动,这样可以帮组它们容易的咬住猎物。
在犬熊属,半熊和恐猫灭绝了以后,锯齿虎和巨頦虎是上新世欧洲的顶级掠食者,植被的变化则是原本独立的草原和树林变的犬牙交错,如有罗盘,锯齿虎和巨頦虎斗能有效地利用这些环紧,后来树林减少以及锯齿虎,豹属的压力(当时虽然也有海德堡人,但是数量太少,基本上就是捡巨頦虎剩饭的),巨頦虎灭绝,而锯齿虎则在四五十万年前在欧洲大陆消失,虽然很北方(北海区域)的锯齿虎一直残延到三万年前。
锯齿虎很长一段时间和西班牙,东欧,中欧,甚至北欧地区和狮子共存,在中国阔齿锯齿虎也和虎共存一段时间,在周口店中更新世的九号,十三号化石点发现了两者共存的证据。锯齿虎和狮子的冲突中应该是弱势的一方,锯齿虎可能会借助林木和树丛避开狮子的骚扰。锯齿虎在中更新世的时候化石就不多了,晚更新世的更是凤毛麟角。
俄罗斯Akhtanizovskaya地区的Tamanian化石点的锯齿虎,亚洲三色豺,和桑氏巨鬣狗
这个化石点的年代110-80万年前,最早在1948年被研究,1957年Vereshchagin介绍了当地的肉食动物,那里面发现亚洲三色豺和桑氏硕鬣狗的下颌骨,此外还有疑似圆锯还有阔齿锯齿虎的材料,里面还介绍比一些小型的肉食动物,比如比狼略小的Canis Tamanensis,这里就不描述了。
亚洲三色豺(Xenocyon Lycaonoides),在Tamanian的早更新世的末期,这里面发现的下颌骨比较大,最大最前面的下颌骨颏孔处于p2的前牙根部的下方,较小的下颌骨颏孔处于p3下方,最小的处于p2后牙根部下方,p2-p4逐渐增高,每一个牙齿比前一个高大概1mm,P4有带状隆凸。
此外该地区还有的鼬科的水獭(Lutra simplicidens),和其他地方的亚种不同的地方在于下裂齿的下后尖较小,下磨牙远中部和下臼齿三尖差不多宽,下磨牙远中部和齿尖和刀片都很发达
这里也发现了很大圆锯锯齿虎的疑似材料,比德国的要大些,下颌骨长度224mm,估测是一个长度330+mm的头骨
猫科的取量,量的是上犬齿的宽度,大概是剑齿虎亚科的明显大于豹属,即便是只有美洲虎或者大雄豹子大小的巨颏虎属(Megantereon)的C种,也超过了狮子了
而豹属里面还是狮子最大,但是欧美洲虎(Panthera gombaszoegensis)的犬齿宽度甚至有和狮子范围重叠的地方,代表他们的犬齿相当的粗壮,接着是豹子和巨猎豹,差不多一个一个级别,再下面是旧大陆山狮,剩下的就是猞猁属,猫属这些小家伙了
其中Vallonnet区域,豹子,锯齿虎,巨猎豹和欧美洲虎共存过,Atapuerca区域狮子和锯齿虎以及一些小型猫科共存过,欧美洲虎也有和巨猎豹+巨颏虎在Valdarno待过
体重表格,欧美洲虎和C型巨颏虎属基本上都是120-170kg之间的体重(虽然可能对C型巨颏虎的上限估测略有高估),但是两者相差不大,没有到狮豹之间的差距,大概是小亚种雌虎和大亚种美洲虎之间的差距吧,巨猎豹比他们都小些,但是明显超过旧大陆山狮(Puma pardoides),狮子和锯齿虎也在一个级别,狮子略大些。法氏三色豺,狼,较大I型猞猁和洞猞猁也是一个级别的。
之间的处于竞争对手位置的有:
早期更新世
1)法氏豺犬和意大利狼(C. etruscus)的竞争很少,但是意大利狼和C. arnensis竞争较多,在没有C. arnensis环境下,前两者都会变小,以利用C. arnensis留下的空缺,在有C. arnensis的情况下,意大利狼会变大,避免和C. arnensis竞争
2)Vallonnet和Untermassfeld地区的C型巨颏虎和欧美洲虎可能有竞争,结果前者被淘汰,而Valdarno-Olivola巨猎豹和豹子的竞争也是前者失败告终,虽然说它们的灭绝时由多种因素造成
中晚更新世
1)狼和亚洲豺,结果豺狗体型缩小,相比之下狐狸受的影响就很少;
2)狮子,锯齿虎,欧美洲虎,豹子,猞猁平均分配资源,虽然最后欧美洲虎和锯齿虎被淘汰,这个估测和环境变化关系更大
上面只是说了犬齿,现在来看一下具体上新世和更新世的肉食动物们喜欢吃什么
表格1:Canidae(犬科), Ursidae(熊科)Procyonidae(浣熊科),Felidae(猫科),Viverridae(灵猫科),Hyaenidae(鬣狗科),Mustelidae(鼬科)
左到右的翻译:素食,杂食,小型动物的肉食,大型动物的肉,昆虫
表格2:Dinofelis(恐猫),Megantereon(巨颏虎),Homotherium(),Smilodon(),Chasmaporthetes(豹鬣狗,一种拥有修长四肢的鬣狗科),Lycaon(三色豺类), Ursus(熊属),Arctodus(短面熊属),Meles(獾属),Nyctereutes(烷熊狗)
里面猫科和鬣狗科,鼬科基本上都其实都在意料之内,唯独那个洞熊符合33%的肉的特征,50%杂食物特征,而食素特征只有17%(按同位素显示则是喜欢基本上吃素),而短面熊居然100%吃素!?(显然不太可能,虽然它确实有能力处理一些素食)这个洞熊的问题,在晚更新世的那篇里面会涉及的,至少罗马尼亚的洞熊食肉性据说也比较高,因此可能有些狮子杀洞熊失败的例子有可能是反过来熊在没有食物的情况下捕杀狮子了。其实主要还是让看看各个动物的下颌骨结构而已。
其实头骨也很重要,看一下头骨的分布
增加了Amphicyonidae-犬熊科,猎猫科-Nimravidae, Thyalcosmilus(袋剑虎),Borthyaenidae(古鬣狗科),肉齿目的creodonta,herpestidae(蒙科)
接下来介绍一下巨猎豹
巨猎豹,顾名思义,比现代的非洲猎豹大,现在非洲猎豹即便是大种群雄性个体也就平均60kg左右,巨猎豹轻松超过这个指标
一下发现的巨猎豹是亚美尼亚地区的,处于欧亚动物群的交际地点
这个生态圈里面已经有比阔齿锯齿虎更大的圆锯锯齿虎,C型巨颏虎,怀特巨颏虎,猞猁,欧亚山狮,欧亚美州虎(panthera onca georgica,Hemmer 2010),更新世鬣狗(P.Perrieri),此外还有各种疣体动物,包括美洲野牛属,马,鹿,羚羊,南方猛犸,犀牛等等,和当时主要的欧洲动物区群区别打算大,当地的环境很多样化,包括了火山灰土,平原,灌木平原,非洲草原,山地和较干燥的半沙漠地区地区
巨猎豹的材料是肢体骨骼,经过比较了一后得出的结果
按肱骨测量结果126kg,用掌骨基酸的结果是按:MCII 92kg,MCIII 98kg, MCIV 102kg, MCV 76kg,不过MCV通常会低估体型,因此意见是100-110kg之间比较可信。
结论是这个个体比较大,应该是雄性个体,较小的标本时雌性
白色的三角形是美洲虎,白色正方形是欧米洲虎,圆圈是奥古斯都美洲虎
旧大陆山狮(Puma pardoides,Owen, 1846),这个动物的化石非常少,头骨几乎没有完整的,根据2010年的一篇依靠鼻孔估测体重的论文里面给出了体重170kg,但是结论显然不可能被接受,前面的一个表格里面估测了45kg,这个显然更接近实际大小,虽然说一些较大的个体体重能更大一些,不过基本上是雪豹到大美洲狮的水平,最大的个体的现代美洲狮子接近吧。
生活年代是上新世到早期更新世的欧亚区域,在西班牙,英国,中欧,巴尔干,亚美尼亚都有发现,下面这些下颌骨就是在西班牙发现的了
下犬齿较长,其实是和损坏有些关系,它和雪豹关系比较近,而雪豹是豹属里面最早和虎一起分化出去的。
西班牙的这个旧大陆山狮股骨比现代豹子(231.1-235.5mm,n=6)要长,也比美洲虎稍微长些(229.8-265.5mm,n=8),在现代美洲狮的范围内(226.3-274.3mm,n=6).在法国发现过长度差不多的股骨,而在德国Untermasfeld的个体大概209-202mm的股骨,假设按比例放大,西班牙和法国的旧大陆山狮可以达到90-100kg之间,股骨的粗壮比例达到了狮子和雪豹的水平,超过了现代的美洲狮和美洲虎
旧大陆山狮子有以下特征:
咬肌窝达到M1的下原尖,三个下颌骨颏孔的位置和现代美洲狮相近,p3前齿冠尖比后面的更突出,p4的下原尖对称/2个齿尖大小相等,m1下原尖比下前尖略大略突出,相当的直挺,肢体骨骼基本上和现代美洲狮相似,除了较为粗壮外
旧大陆山狮子和雪豹相似的特征:
下颌骨坚固,下颌骨颏孔的位置和数量,咬肌窝达到M1的下原尖,p3有一个圆形的前柱,p4的下原尖对称/2个齿尖大小相等
和雪豹不同的特征包括:m1下原尖比下前尖略大略突出,下原尖前段有一个小突状物,下颌骨粗壮程度,较短和粗壮的股骨
虽然有一些三百多万年前的疑似美洲虎的豹属化石(Barry, 来源于非洲),虽然那个化石最后被认为是原始的狮类(Turner),但是早期狮类的上裂齿p3-p4确实和欧美洲虎很相似,一些来自Siwaliks印度的花是则被认为是最早的美洲虎类(turner),美洲虎类于上新世晚期进入美洲。所有的美洲虎都是一个种的,但是有不同的亚种,例如欧洲的欧美洲虎(P.onca gombaszoegensis)和P.onca toscana,已经巨大的美洲奥古斯都美洲虎P.onca anugista。
以下介绍一的化石来源是土耳其和黑海之间的古亚美尼亚地区,和前面的巨猎豹来自一个同国家,但是不是同化石点,巨猎豹比它早90万年前,不过和巨猎豹在一起发现的也有亚美洲虎(也可能是欧美洲虎的疑似材料)
环境基本上和前面巨猎豹的栖息地是一样的,作者对已经确认种类的素食动物总数统计的结果是:
犀牛类0.2%,原大角鹿29.1%,马属27%,猛犸1.9%, 土拨鼠2%,牛类8.5%
发现的欧美洲虎下颌骨;按照下裂齿联合长度(Valkenburgh),大概估测结果100-110kg,这样算至少是很大的巴西帕塔地带的大亚种现代美洲虎雄兽的极大个体水平。不过这个标本的牙齿比例较小,实际体重应该要大些,Hemmer提出用下裂齿估测现代美洲低估了体重接近20%,因此作者认为真是的体重应该是130kg,属于很大的雌性或者中等大小的雄性。当然其实还有一些欧美洲更大,下面会提到的。
Akhalkalaki的标本德年代大概在90万~80万年前
牙齿特点:p3比例值(77.5)在欧美洲虎的变异范围内(英国 71.5-78, 意大利71.5-78),但是比德国(68-74.5)高,比古虎类(P.tigris "palaeosinensis", P.tigris "oxygnatha", ,70.25)和比亚洲狮子更大(63.5-65.5),宽度比例则和德国的欧美洲虎差不多,比其他地区的美洲虎稍宽,但是还是在变异范围内,和史前狮虎比较接近,Akhalkalaki的标本的p3的另外一个特征是前端非常的狭窄,和德国的比例值比较接近(87 vs79-91.5),值数超过意大利的,明显超过狮虎
M1的长度相对很大,比例值109.5,但是在欧美洲虎的范围内(100-112.5),超过了虎(97.5-105.5)和亚洲狮(99.5-104),古狮(97.5-105.5)
牙齿测量表格,上面也有奥古斯都美洲虎按牙齿长度估测,P3-M1有66mm,表格上的最大个体可能比这个欧美洲虎个体大10+%,不过欧美洲虎肯定有更大
欧美洲虎个体在对抗生态地位中低于圆锯锯齿虎属,其余的情况下至少可以和中小数目的亚洲三色豺和桑氏硕鬣狗对抗
现在看一下正宗欧洲地区的欧美洲虎,这是一个Tegelen荷兰的标本,也是最早的欧美洲虎之一,更新世早期(170~240万年)的一个化石点
标本号NGM103142,左侧下颌骨
特点:下裂齿的p3几乎没有前齿尖(anterior cusp),上原尖很大,末端后齿尖也很小但是很明显,P4上原尖很大,向后倾斜,前齿尖很明显,带状隆凸部分清晰可见,但是M1残缺,下前尖平滑,没有多出的突起,下颌骨颏孔共有两个,较小的那个位置处于牙间隙下面,咬肌窝相当的深,紧接处于于M1下后方,p4和M1有明显的重叠,牙齿挤在了一起,但是p3和P4之间距离正常,根据牙齿磨损情况看,这个是一个体成熟的成年个体,大概同等于7-9岁的狮子差不多(Smuts)
另外一具标本NGM 102738,
特点:下犬齿完整,相当的大,p3很大,所有P4的齿尖都很大,左下颌骨p4前齿尖非常的明显,有下颌骨的p4前齿尖也很大,但是比较圆滑,上原尖笔直,咬肌窝相当的深,P4和M1有较小的重叠,和前面的标本不同的是P3有很明显的前齿尖,前面的那个比标本没有这个特点是因为磨损所致(不过欧美洲虎的下裂齿p3变动很大,不能作为鉴定材料),M1上面有明显带状隆凸和下前尖
测量表格:NGM 102738比NGM103142更大,很可能是它是雄性,后者是雌性
同样也可以看出这个NGM 102738标本明显超过了前面的亚美尼亚的那个欧美洲虎,但是按M1大小看L Escale的和德国Untermassfeld和它差不多,甚至有比它还要略大的个体
而且这个P3-M1的长度可达75mm,也超过楼上表格里面的奥古斯都美洲虎的大小,另外那些AM#标本都是阿姆斯特丹博物馆的欧亚草原狮/古狮什么的,所以明显超过欧美洲虎。
这个荷兰的欧美洲虎的大小在欧美洲虎内部的比例,显然德国比它要大很多,但是这个荷兰标本的P4很长,所以实际大小比看M1要更大些
下图是荷兰欧美洲虎和南非的狮子的比例
下面是所有欧美洲虎和现代南非的狮子的比例,可以看出最大的欧美洲虎大概有一个大雌狮或者小雄狮的水平,而且美洲虎前面说了,M1和P4比例都比较小,所以这个标本估测体重大概要至少比前面那个亚美尼亚的欧美洲虎(100-130kg)大30+%,重量大概(130-175kg),而德国的标本(140-180kg也是可能的),欧美洲虎整体上有大型岛屿虎的雄兽的水平,甚至有些小型的狮子亚种的水准,肯定是可以和奥古斯都美洲虎一拼高下的。
国家的顺序是(country),乔治亚,保加利亚,荷兰,希腊,意大利,希腊,德国,乔治亚,德国
前面说到的akhalkalaki的标本可以说是最晚的欧美洲虎了
鉴定特点包括:上裂齿P4上面有Ectoparastyle,还有笔直的前端以及弯曲的后尖,上裂齿P3的前后齿尖大小不一样,变化很大,但是带状隆凸很明显,上犬齿很粗壮,矢状嵴很发大,
俄罗斯的洞狮,以及记载的一些欧美洲虎和豹子的数据,颈背冠和枕基也是鉴别特征之一,门齿排列成直线,上门齿I3最大,有一个椭圆的眼窝下孔,鼻腔开口很宽,鼻腔骨衍生到了头盖骨/上颌骨缝合线的地方甚至衍生到其之后。具体的图片,那个《大猫和其史前化石亲戚》里面有介绍,英雄王有着本书
巨颏虎属(Megantereon)的介绍大管家已经详细的说过了,因此不再进行复述,只是详细的描述一下法国地区的完整标本肢体部分的大小
法国Seneze的一个几乎完整的C型巨颏虎(M.cultridens)的骨架,它的年代鉴定是上新世末期,
按骨骼公式估测这个材料的头骨和肢体并不算特别大,基本上是肱骨平均110kg,尺骨107.5kg,股骨101.3kg,胫骨105.7kg,
假设活的时候肩高73-85厘米左右,骨骼体长130厘米左右,活体体长152-167厘米,估测体重按美洲虎估测至少有100kg左右,(按等比例放大,按狮虎等比例算就小的多了,88kg,不过它肢体比美洲虎都大,但它比狮子,虎,豹子和美洲虎粗壮),加、如果取上限的话,可以到中小型的雌孟虎的体形,考虑到比较粗壮体重大概120-140kg,取下限也有100kg左右。前面的表格也说到了这种巨颏虎肯定能有超过100kg的个体。甚至说160kg都有可能。
脊椎骨的测量数据
颈椎骨相对很长,显得很纤细,但是实际上最大长度(包括两翼)直达现代狮虎的下限。像锯齿虎一样,颈部活动能力很强
颈椎骨比较:
A-E是C型巨颏虎,F-J是致命刃齿虎,K-O是狮子,P-T是豹子
整体上神经枢是类似于刃齿虎椭圆的,而不是锯齿虎属的圆形的
脊椎骨的比较
C型巨颏虎的骨盆
肩胛骨和前肢骨骼的大小,具体看图片的介绍
C型巨颏虎的肩胛骨长度217.8mm和狮子的范围重叠(雄210.2-288.4mm, 雌175.7-242.1mm,n=17),和孟虎(雄,190.7-241.7mm, 雌199.1-241.3mm,可能取样雄性没很大),西虎的肩胛骨也很大(270.5-284.8mm,n=3),,C型巨颏虎的肩胛骨长度超过美洲虎(152.1-172.6, n=8), 和豹子(123.2-170.3mm N=15), 但是远不如致命刃齿虎(266-358mm)
巩固长度262.5mm,在狮虎的下限范围内,但是远比美洲虎大(195.8-238.7mm, n=8),比豹子和美洲狮就更大的多了(豹,n=16,176.4-222.6,美洲狮 n=6 187.4-231.9),但是远小于刃齿虎(353.4-422mm)
粗壮程度:肱骨粗细最小周长/全长
C型巨颏虎(0.356)超过狮子(0.318),美洲虎(0.322),虎(0.303),Neofelis(0.298),美洲狮(0.291),纤细刃齿虎(0.330),致命刃齿虎(0.337),仅仅逊色于最粗壮的毁灭刃齿虎(0.401mm)
肱骨头的粗壮程度也仅仅逊色于毁灭刃齿虎。
尺骨鹰嘴的比例,尺骨的杠杆效力显示是鹰嘴占尺骨比例最大最佳,可以迅速的从伏击的姿势跃起,C型巨颏虎和毁灭刃齿虎最强,另外参考棕熊挖洞的,尺骨鹰嘴也很强,C型巨颏虎和毁灭刃齿虎牵制收缩,拉,扯,拔倒猎物的能力也比较好,它的桡骨和肱骨的比例和恐猫差不多,超过美洲虎和刃齿虎,类似于半伏击猎手,即能伏击也能短距离的追击,虽然追击能力还是不如隐剑齿虎和豹子
后肢骨胳的比较
股骨长度286mm,比那个旧大陆山狮要长了,明显超过美洲虎(230-265.5)和豹子(201-255.3)
,达到了狮虎的下限范围,股骨粗壮程度(最小周长/全长)有0.289,明显超过美洲虎(0.263),豹子(0.246),狮子(0.271),虎(0.252),新猫类(0.255),美洲狮(0.234)和纤细刃齿虎(0.263),和指明刃齿虎持平(0.288),不过还是低于毁灭刃齿虎(0.324),致命剑齿虎有423.5mm, 毁灭刃齿虎的股骨引用的是PC的2个个体最大有395mm的,其实肯定能找到更大的标本。
骨盆:形态强壮,按比例宽度超过刃齿虎和现代大猫
长度260.8mm, 狮子(245-333mm),虎(256.5-345.3mm),豹子(165-218.2mm),美洲虎(200-227mm),致命刃齿虎(283-368mm),锯齿虎(322mm)
胫骨:粗壮程度(0.308),远远超过狮虎(0.275),美洲虎(0.281),豹子(0.240),美洲狮(0.236),纤细刃齿虎(0.290),但是不如巨形剑齿虎(0.323),致命刃齿虎(0.323)和毁灭刃齿虎(0.378!)
PS:虽然说per-C的体重攻势估测的结果感觉刃齿虎的后肢比较弱,其实并不是这样,粗壮程度显然和前肢一样出色。
肩胛骨占的比例高大前肢的38.6%,肢体的杠杆效力甚至好过刃齿虎属,这个英雄王原来已经解释过了,但是它的后肢比例看,跳跃能力较差(跳跃能力最好的是前肢相对后肢比例最小的雪豹和美洲狮),桡骨和肱骨的比例前面已经说过了,整体上看,C型巨颏虎最接近现代猫科中的美洲豹和史前猫科中的刃齿虎。
C型巨颏虎是林栖型的动物,但是居住的不是浓密的树林,也有可能进入开阔地(当时环境是林原交错)在开阔地捕猎的时候它会尽量选凹凸不平的地形作掩护,接近猎物接着短距离出击。它和其他剑齿虎类一样,利用强壮的前肢压制猎物,精确的杀伤猎物的要害,接着借助头颈肌肉辅助咬入猎物。
C型巨颏虎最可能是有蹄类,和致命刃齿虎一样,不过猎物体形相对小些而已
PS:其实除了血刃刃齿虎之外其他的都不怎么猎杀较大的厚皮动物
包括猴类,猛犸,各种鹿,野猪,肉食动物则有豹鬣狗,猎豹,锯齿虎,巨颏虎,熊,狐狸,灵猫类,狼/犬类,硕鬣狗
根据同位元素研究,显示巨颏虎喜欢伏击林中的原大角鹿类,而阔齿锯齿虎喜欢吃大型野牛和幼年猛犸,虽然大角鹿类的角会给猎手咬后颈造成不便,也许巨颏虎会选择角较小的雌兽和幼兽
阔齿锯齿虎和巨颏虎的生态位置也不是完全分开的,它俩也都喜欢吃马类动物,
C型巨颌虎长开大嘴巴的样子
现在浅谈一下怀特巨颏虎(Megantereon whitei),它比C型巨颌虎要小很多,只有一个豹子那么大,但是它更加的先进,取代了前者的生态地位,它在上新世末期从非洲演化,并在早更新世进入欧洲,在意大利就发现过它的化石,这是化石和测量数据,意大利的标本是AGR系列的,看地图
其实可以发现看掌骨测量数据,SE311并不是最大的,而且怀特巨颏虎头骨长度和体形虽然明显比C型巨颏虎小,掌骨却不算太小
下面是头骨的对比,a是C型巨颏虎,c,b是怀特,c就是AGR标本的头骨了,a明显比c大的多,有兴趣的可以拿它和前面的SE311比一下。
化石AGR系列原状态
C型巨颏虎的起源是美洲,接着在早上新世进入欧亚非,在早期更新世的欧洲灭绝,在中国则一直延续到了中更新世,M.falconeri是最大的巨颏虎,用C型演化而来,起源是印度,怀特种的起源是非洲,接着进入了亚美尼亚,土耳其地带,最后进入西班牙,希腊,意大利,希腊发现的个体的变异范围就是属于,怀特种像所有的剑齿虎亚科一样,留下大量的食物给食腐动物,这包括了早期人属,在怀特种进入西班牙,土耳其和亚美尼牙区域的时候,化石点也出现早期的人类(palmqvist 1995, Martinez-Navarro)
显然是印度的最大,印度的Megantereon falconeri.最大的巨颏虎,看体形可能不会比大亚洲狮虎的雌兽小,甚至有东非成年雄狮的中小个体差不多
Megantereon falconeri的下裂齿p4,下裂齿M1长度完全可以和前面发的欧美洲虎匹敌,其上裂齿P4更是非常的大。意大利的比法国的略大。非洲的最小,估计是怀特种
Venta Micena西班牙的巨颏虎同在的种群包括,这个是晚上新世转化早更新世的革命
土著:南方猛犸,阔齿锯齿虎,意大利双角犀,意大利熊,
亚洲来客:主要是一些中型的有蹄动物,还有鹿类,原大角鹿
非洲来客:桑氏巨鬣狗,法氏三色豺,古河马,怀特巨颏虎
西班牙发现的下颌骨
世界个体的巨颏虎属的对比分析
白圆是这次的版本,黑圆是非洲标本,都是怀特种,黑菱形是欧洲,白菱形是印度,白三角是北美,黑三角是中国,印度的是F型,明显最大,北美和非洲的都似乎是C型,中国的是自己几个种
最后谈一下Megantereon whitei和M. cultridens的区别
大管家很早就总结了,引用他的话
“Turner等人认为Megantereon whitei实际上就是M. cultridens的同物异名.所谓的非常小,只是由于性别不同所产生的误差.而p3是退化中的牙齿,很难根据其大小来鉴定种.但是,后来Palmquist等人分析了雌雄两性的狮子,雌雄两性的豹子,与两种巨颏虎之间的差异,断定Megantereon whitei并不是雌性的M. cultridens.”
这里面给一些详细的比较。
头骨大小,很直观,W型的牙齿更退化一些
结果:中国的尼河湾种和C型很接近,但是W型差距比较明显,差异程度超过了现代的猫科里面的雌雄差距,少数差距小于现代猫科雌雄的仅仅是上犬齿的长度和宽度。
但是裂齿和前臼齿的差距就很大了,P4在W型里面的相对大小比C型更小,明显超过现代猫科。
显然,M型和C型之间差距大于现代猫科雌雄
而下裂齿的下原尖和下前尖的分析也显示同样的结果
差距超过雌雄豹子和雌雄狮子之间
两者头骨的区别,而且放大W型的头骨到C型的大小,头骨抗反作用力的能力和机械效率都比C型高,所以如果M型打算长大,它显然应该保留自己头骨的形态而不是变成C型的样子,看上面那个表格,很容易明白的,显然W型的抗反作用力能力(Y轴)高于同大小的C型头骨,MAT是枕骨到下颌骨冠突的距离,大概就是和大小成正比了。
头骨的一些特点包括了
1)上犬齿薄而长,没有锯齿
2)门齿很厚实,帮助撕肉,3)上裂齿的上前尖退化,和猎豹很接近,显示它们只特化吃肉,尽量不接触骨头,下颌骨冠突减少,头骨宽度减少(毁灭刃齿虎也有这样的特征),咬肌肉减少,位置变化,增加下裂齿的咬力,头颈部肌肉辅助咬力。
W型取代C型的原因是相对更加进化的头骨可以让W型以较小的体形完成捕杀同样大的猎物,W型的嘴巴可以张的更大,当然这样退化下颌骨冠突也导致咬力减少,为了弥补这点,W型把不需要用类处理食物的一些牙齿都退化,减短头骨和下颌骨的长度,一边增加杠杆效力,达到增加咬力的效果。
巨颏虎属在很大程度上帮助了人类和鬣狗的兴旺,尤其是在欧洲较冷天的天气,肉不会腐烂的那么快,给人类提供的肉剩余就较多,早期海德堡人和巨颏虎属共存。
(本文为化石网论坛精华帖,由网友鸿穹神皇上帝提供,仅供大家学习参考。编辑:常君滢)
2022-11-29