-

沧海之牙(一)-沃土催芽
每年都有不少人因冲浪死于鲨口 , 其中澳大利亚海域是全球大白鲨伤人最频繁的区域 , 世界各地发生的鲨鱼攻击人类的事件中,公牛鲨和鼬鲨袭击人类的次数更不亚于大白鲨。假设置身于第三纪的海洋,你会发现第三纪
每年都有不少人因冲浪死于鲨口 , 其中澳大利亚海域是全球大白鲨伤人最频繁的区域 , 世界各地发生的鲨鱼攻击人类的事件中,公牛鲨和鼬鲨袭击人类的次数更不亚于大白鲨。假设置身于第三纪的海洋,你会发现第三纪的海洋会比现在的海洋可怕百倍,更能符合“地狱水族馆”的称号 ,当时的海洋不是只有大型鲨类在游弋,还可以看到掠食性的抹香鲸类捕食的情景 ,也许可以看到大型史前抹香鲸如迅捷鲸Ontocetus oxymycterus或斯卡尔鲸属种Scaldicetus caretti和巨齿鲨搏斗的壮观情景 , 后来的黑鲸类兴起更使第三纪海洋变得更精彩 。就算是浅海及海岸 , 海牛及岸上的海象等鳍足类,海陆两栖的鳄类的多样性都使行人望而却步。 幻想在当时的海洋冲浪,恐怕自己突然会掉进一个血盘大口!
于<<沧海之牙>>中会介绍晚侏罗纪至第四纪鲨类的种类及演化,并会详实探讨鲨类的分布变化 , 本贴会介绍的鲨类是包括虎鲨目,扁鲨目,锯鲨目,须鲨目,六鳃鲨目,笠鳞鲛目,角鲨目,鼠鲨目及真鲨目。
绞口鲨科(Ginglymostomatidae)有非常长久的历史,现在的绞口鲨科(Ginglymostomatidae)包括铰口鲨(Ginglymostoma cirratum), 短尾护士鲨 (Pseudoginglymostoma brevicaudatum)及光鳞鲨(Nebrius ferrugineus)。此外还有不少已灭绝的属种包括副绞口鲨(Paraginglymostoma) , 关于副绞口鲨Paraginglymostoma的地位是一个问题 , 副绞口鲨Paraginglymostoma被认为可能是现存的绞口鲨科成员的姐妹群。副绞口鲨Paraginglymostoma是一种小型鲨类并局限于马达加斯加岛及东非海岸的一小部分 , 绞口鲨科仅存的成员移动性要较其他须鲨目成员来得高。始新世西欧及中新世非洲也存有另一种史前绞口鲨科成员原铰口鲨Protoginglymostoma ypresiensis 。
史前的绞口鲨科成员还有生存于古新世的非洲 , 始新世北美的Delpitoscyllium , Delpitoscyllium仅只有Delpitoscyllium africanum一种 , Delpitoscyllium africanum曾被划作铰口鲨属 , 但牙齿上的不同使它自成一属 , Delpitoscyllium africanum牙齿主齿尖有二个小齿尖 , 牙齿唇舌面侧部扁平压缩 ,这点有别于光鳞鲨属及铰口鲨属 ,于渐新世地层再没有任何Delpitoscyllium africanum牙齿化石发现。
Plicatoscyllium 是生活于晚白垩纪时期坎帕阶(Campanian)至马斯特里赫特期(Maastrichtian)北美及摩洛哥的绞口鲨科(Ginglymostomatidae)成员 , 属种包括生活于北美马里兰州及德克萨斯州的P. antiquum 及 P. derameei(P. derameei和Ginglymostoma lehneri是同物异名的) , 摩洛哥的Plicatoscyllium gharbii 及P. youssoufiaense , 还有分布广泛的Plicatoscyllium minutum , Plicatoscyllium minutum于坎帕阶(Campanian)至马斯特里赫特期(Maastrichtian)已分布于西非,欧洲,北美东部 , 西印度及德萨斯州 。Plicatoscyllium minutum牙齿很独特 , 有一个主齿尖并伴随四个高的侧齿尖 , 这些侧齿尖都很宽阔及尖锐 , 也有较高的牙根 。
于英国晚白垩纪地层曾发现了一类已灭绝的绞口鲨科成员部分椎骨化石 , 这类鲨鱼是Cantioscyllium , Cantioscyllium属牙齿和一般的绞口鲨相似 , 有着较低且粗大 , 向舌的齿冠 , 齿根向唇的位置呈V型 , 有较圆及细小的侧齿尖 , 于齿冠及齿根中央各存有一个孔 , 前列牙齿有较直而尖的齿冠 , 齿冠侧部位置膨大, Cantioscyllium牙齿适合抓捕猎物 。
Cantioscyllium于森诺曼阶至土仑阶(Turonian)的欧洲西部及北美内海繁盛起来 , 于德克萨斯州及肯特立的森诺曼阶(Cenomanian)至科尼亚克阶(Coniacian)地层,甚至连马里兰州及德克萨斯州马斯特里赫特期(Maastrichtian)地层也有Cantioscyllium meyeri牙齿化石发现 , 乌兹别克斯坦土仑阶(Turonian)地层发现了Cantioscyllium nessovi , 于北卡罗莱纳州坎佩尼阶(Campanian)地层也发现了Cantioscyllium decipiens牙齿化石 , 法国森诺曼阶至土仑阶(Turonian)也有相似种Cantioscyllium cf. decipiens的牙齿化石。 此外于2002年西欧晚侏罗纪地层发现了Cantioscyllium未定种的牙齿化石,显示Cantioscyllium最早该于晚侏罗纪已存在。于西班牙东北部Teruel省的白垩纪前期巴列姆阶(Barremian)出土了达十三种软骨鱼类的牙齿化石 , 其中有四种是新种包括史前鳐类"Rhinobatos" incidens , Celtipristis herreroi , 虎鲨属Heterodontus carerens及Cantioscyllium alhaulfi , Cantioscyllium alhaulfi就是刚发现的Cantioscyllium属新种。于当地也发现了其他软骨鱼类包括Cretolamna sp., Carcharias sp., Onchopristis sp. , Protolamna sp. cf. P. sokolovi, Protoscyliorhinus lamaudi, Spathobatis halter, "Rhinobatos" picteti, Pseudohypolophus mcnultyi 及Onchopristis dunklei 。
虎鲨目(Heterodontiformes)又称为异齿鲛目,是板鳃亚纲中的一个比较小的目,现存只有一个虎鲨科及一个虎鲨属,包括现存的九种比较小的,生活于热带及亚热带深海底及海岸的鲨鱼。这些鲨鱼的长度在50-150厘米之间。它们一般以生活在海底的海星、贝壳、蟹、虾等为食。除了瓜氏虎鲨(H. quoyi)是生活于秘鲁海岸及加拉帕弋斯群岛邻近外,虎鲨目成员大多数于太平洋板块及海岛邻近都是没有踪影的 。现存虎鲨目成员大多是分布于东太平洋 , 于东非,阿拉伯半岛,亚洲东南部至日本及澳洲都可发现其踪影 , 佛氏虎鲨H. francisci及墨西哥虎鲨H. mexicansus局限于北美西部。不过最早的虎鲨目是来自早侏罗纪的德国北部 , 于侏罗纪早期至中期的欧洲特提斯海西部是存有许多岛屿 , 当时环绕着特提斯海东北部的印尼-马来西亚地区 , 欧洲中央及西部的北非 , 东南部的南极及澳洲的气候属于亚热带至热带 , 在北极盆地气候被认为是和暖的。侏罗纪的虎鲨目和现存的种类非常相似并且是同属的 , 虎鲨属扩散至古大陆(Pangaea)东部的海岸地带已有相当长久的历史 , 是古大陆分裂做成了今天的虎鲨属的分布。于南美巴塔哥尼亚及阿根廷白垩纪地层 , 美国乔治亚州始新世地层 , 维吉尼亚中新世地层也发现的虎鲨属种化石显示它们曾于大西洋也有分布。
关于虎鲨科早期历史资料仅来自零散的牙齿化石发现 , 于英国侏罗纪中期及法国侏罗纪晚期地层就发现了原虎鲨属Proheterodontus牙齿化石 , 但有一种虎鲨科成员较原虎鲨属Proheterodontus分布还要广泛,于英国,比利时及德国南部侏罗纪早期至晚期地层就发现了副异齿鲨属Paracestracion化石 , 侏罗纪早期至中期出土的大多是牙齿化石 ,于侏罗纪晚期地层也有铰接的骨骼化石发现 。于德国南部的Nusplingen及Solnhofen石灰岩上就发现了副异齿鲨属---镰状副异齿鲨Paracestracion falcifer 的成年及幼体化石样本 , 副异齿鲨属Paracestracion的腹鳍在第一背鳍之下,两个背鳍都有鳍棘 , 于镰状副异齿鲨Paracestracion falcifer的背鳍棘有部分是存有血管,这点和现存的虎鲨属是相反的。副异齿鲨属Paracestracion幼体有梳状的牙齿 , 没有像臼齿般的侧齿 , 相反现存的虎鲨属有臼齿般的侧齿 , 幼副异齿鲨属Paracestracion及现存的虎鲨属侧齿都没有充足的有机物但成年的副异齿鲨属Paracestracion侧齿却有足够的有机物 , 发现的镰状副异齿鲨Paracestracion falcifer幼体及成体长度分别长约25厘米及42厘米。
近来于德国南部启莫里奇阶(Kimmeridgian)晚期地层也发现了副异齿鲨属Paracestracion的新种---维奥尔副异齿鲨Paracestracion viohli , 这维奥尔副异齿鲨Paracestracion viohli个体是属于一个亚成年个体 , 没有臼齿形的侧齿 , 维奥尔副异齿鲨Paracestracion viohli牙齿都有着其他副异齿鲨属种的特征, 牙齿形态及大小都和成年副异齿鲨Paracestracion属种及虎鲨Heterodontus属种相似 ,但不同的是维奥尔副异齿鲨Paracestracion viohli牙齿齿冠表面有纹 , 它的牙齿齿根血管系统也和其他副异齿鲨属种不同。由于发现的维奥尔副异齿鲨Paracestracion viohli个体牙齿没有像镰状副异齿鲨Paracestracion falcifer幼体般呈梳状的牙齿 , 也没有臼齿状的侧齿,因此化石个体是不属于幼体且被认为是接近成年的 , 该化石个体长度估计长约30厘米。
原虎鲨属Proheterodontus及副异齿鲨属Paracestracion于侏罗纪至白垩纪交界便告灭绝, 但虎鲨属却一直延存下去并于白垩纪及第三纪时期变得更具多样性。最早的虎鲨属是一亿五千万年前的Heterodontus zitteli及晚侏罗纪启莫里阶(Kimmeridgean)法国的Heterodontus semirugosus。Heterodontus zitteli和副异齿鲨属Paracestracion外型相似,但是二者不同的是,Heterodontus zitteli腹鳍位置是在第一背鳍后方。史前虎鲨属于欧洲 , 非洲 , 澳洲及美洲的白垩纪地层都有化石发现包括白垩纪早期波兰的Heterodontus polonicus , 白垩纪法国及英国的Heterodontus canaliculatus , 晚白垩纪科尼亚克阶(Coniacian)至坎帕阶(Campanian)德克萨斯州的H. cf canaliculatus , 阿尔布阶(Albian)英国及法国的H. upnikensis, 晚白垩纪坎帕阶(Campanian)密西西比州的H. rugosus , 晚白垩纪坎帕阶(Campanian)西欧的H. havreensis , 晚白垩纪马斯特里赫特阶(maastrichtian) 美国新泽西州的H. creamridgensis 及德克萨斯州的H. granti , 古新世至始新世维吉尼亚的H. lerichei ,早始新世密西西比州的H. sowasheense , 始新世北卡罗莱纳州的H. elongates , 晚始新世乔治亚洲的H. pineti ,始新世中期乌兹别克及英国的H. vincenti,始新世中期至晚期英国及美国的H. woodwardi , 渐新世北卡罗莱纳州的H. jaqnefirdae , 渐新世至上新世澳洲的H. cainozoicus 等等 , 现存的日本虎鲨H. japonicus也于挂川上新世地层有化石记录。于始新世之后 ,虎鲨属于大西洋西部便开始消失 , H. jaqnefirdae一直坚持至渐新世时期,但于当地中新世沉积层再没有虎鲨属的化石记录 , 但于东太平洋虎鲨属还是依然的普遍繁盛。值得一说的是白垩纪森诺曼阶(Cenomanian)哈萨克也有虎鲨科中的伪虎鲨Pseudoheterodontus polydictus , 虎鲨科还有中新世北美的Strongyliscus , 不过资料太不流通。
扁鲨科仅有一属十六种 ,大多生活于气候温和的海岸及热带地域半海底上层, 尽管扁鲨科也有些体型较大的属种,但它们倾向底栖的 , 扁鲨科是不会出没于海岛旁的水域。最早的扁鲨科成员是来自晚侏罗纪的德国 , 于白垩纪及新生代大幅度扩充 , 可能来自早期超大陆的分裂从而做成地理上的分隔 , 它们于盘古大陆(Pangaean)旧海洋中大幅度扩散并做成现在的分布。
侏罗纪的扁鲨科有伪莱茵鲛属Pseudorhina , 属种包括Pseudorhina alifera 及Pseudorhina acanthoderma二种, 对这些侏罗纪的属种认知大多数仅来自离散的牙齿化石 , 根据形态分析伪莱茵鲛Pseudorhina是被认为和扁鲨属都是单源的 , 彼此有着显著形状的舌基软骨及牙根结构 , 透过和现存的扁鲨属的骨骼形态比较 , 显示侏罗纪的扁鲨类形态和现存的扁鲨已相似得很 , 但于一些特征如眼眶结构 , 眶后突 , 筛骨区的孔位置等却有所不同。伪莱茵鲛Pseudorhina也有着现存的扁鲨属数个起源特征 , 例如有三角形及膨大的胸鳍叶,其他的特征不在此详说了。
扁鲨属可能于晚侏罗纪已存在于地球上 , 英国的侏罗纪启莫里奇阶(Kimmeridgian)地层发现的 Squatina frequens该是最早的扁鲨属种,不过Squatina frequens归属尚存有疑问。于白垩纪时期坎帕阶(Campanian)至麦斯特里希特阶(Maastrichtian)的欧洲及北美就生存了一种史前扁鲨属种凯斯扁鲨Squatina hassei , 凯斯扁鲨Squatina hassei牙齿只有5 mm长, 牙齿两旁还长有倾斜短小的小齿尖 , 凯斯扁鲨Squatina hassei是一种细小的扁鲨类 , 于晚白垩纪的西大西洋也有凯斯扁鲨Squatina hassei的化石记录。阿尔布阶(Albian)英国及法国,森诺曼阶(Cenomanian)德国的Squatina cranei , 它的牙齿和现存的扁鲨(Squatina squatina) 相似, Squatina cranei也有部分骨骼化石被发现; Squatina decipiens的生存年代及地域和Squatina cranei有所重叠 , 但是哈萨克土仑阶(Turonian)地层也有Squatina decipiens的牙齿化石发现,此外阿尔布阶(Albian)英国及法国也存有另一种扁鲨属种Squatina mulleri ,晚白垩纪坎帕阶(Campanian)德国的Squatina baumbergensis 则有完整的化石发现 。至于古新世时期至始新世时期的扁鲨属种有Squatina prima , 它的化石于摩洛哥 , 美国切诺皮克,比利时及英国都有发现 , 化石属于古新世(达宁期Danian至塔内提阶Thanetian)至始新世伊普雷斯阶(Ypresian) , Squatina prima有纤弱的齿冠及较低的侧翼。于渐新世至中新世有一种分布广泛的扁鲨属种Squatina subserrata , 最初被发现于中新世的奥地利 , 后来于北美马里兰州也发现了Squatina subserrata的牙齿化石 , 它的牙齿要比古新世至始新世的Squatina prima牙齿来的结实 。第三纪的扁鲨属除了Squatina subserrata ,还有始新世比利时的Squatina crassa ,渐新世德国的Squatina angeloides , 加利褔亚也发现了中新世的扁鲨属种Squatina lericheii ,美国约克登(YorkTown)上新世地层也发现了杜氏扁鲨(Squatina dumeril)的亲近种S. aff dumeril 牙齿化石 , 上新世智利也有另一个史前扁鲨属种Squatina biforis 化石发现。 此外日本也发现了未定种的扁鲨属牙齿化石, 该未定种牙齿化石是属于渐新世时期的。现存的扁鲨属种大概于中新世时期已存在 , 像非洲扁鲨(Squatina africana)于南非中新世至上新世地层已有化石记录 , 加州扁鲨(Squatina californica)于圣地牙哥上新世地层也有化石记录 , 上述的杜氏扁鲨(Squatina dumeril)亲近种也于美国约克登上新世地层被发现。
锯鲨目有一科两属 , 六鳃锯鲨属仅只有瓦氏六鳃锯鲨Pliotrema warreni一种 , 生活于南非 , 莫桑比克南部及马达加斯加岛东南部气候温和的浅海及半深海区上部, 锯鲨属则有四个已确认的属种及四个未定种。较大的属种像长吻锯鲨(Pristiophorus cirratus), 日本锯鲨(Pristiophorus japonicus)及未定种的P. sp. A(东澳大利亚锯鲨)是生活于西太平洋温和的浅海 , 较细小的属种如巴哈马锯鲨(Pristiophorus schroederi), P. sp. B(热带澳大利亚锯鲨) , 及P. sp C(菲律宾锯鲨)则于北大西洋西部及太平洋中心的西部有广泛但不相连的分布。短吻锯鲨(Pristiophorus nudipinnis)则生活于澳洲南部 , 印度洋西部则有P. sp. D(侏儒锯鲨) , 属种间的相互关系是备受研究中。锯鲨属于晚古新世的摩洛哥, 晚始新世俄勒冈州, 渐新世比利时,荷兰及加利褔亚, 晚渐新世的日本,中新世德国西南部 , 葡萄牙,法国南部及上新世的新西兰都有化石记录 , 锯鲨类最初分布于广阔的特提斯海 , 于渐新世时期则存活于东太平洋 , 至现在则灭绝于东太平洋 , 于大西洋还有残存的种类, 在澳洲则表现出高度的多样性。
关于史前锯鲨目成员的资料没多少可以备受陈述的, 最古老的锯鲨科成员化石是晚白垩纪黎巴嫩的原锯鲨Propristiophorus , 它的外型已和现存的锯鲨类相似。此外始新世至更新世早期新西兰有Ikamauius ensifer , 史前的锯鲨科成员还有Rhaibodus , Rhaibodus属的具体资料完全不详 ; 至于锯鲨属Pristiophorus的史前属种包括古新世摩洛哥的Pristiophorus palaeocenicus , 渐新世日本的Pristiophorus lineatus, 渐新世比利时的Pristiophorus rupeliensis , 始新世至更新世智利 , 新西兰及澳洲的长寿种Pristiophorus lanceolatus , 上新世新西兰的Pristiophorus napierensis, 中新世奥地利,法国,荷兰及德国的Pristiophorus suevicus 。现存的巴哈马锯鲨(Pristiophorus schroederi)于芬兰中新世地层 , 比利时上新世地层已有化石记录 ,琉球群岛上新世地层也有日本锯鲨(Pristiophorus japonicus)的化石记录 ,南非中新世至上新世地层已有瓦氏六鳃锯鲨Pliotrema warreni的化石记录。
须鲨目Orectolobiformes是从史前的原始鼠鲨目成员分化过来的 , 一共有两个亚目,分别是喉须鲨亚目及须鲨亚目 ,但以下介绍的鲨类都是于分类学上存有疑问的。
最早期的须鲨目成员是早侏罗纪锡内穆阶(Sinemurian)至普连斯巴奇阶(Pliensbachian)欧洲西北部的Agaleus dorsetensis , Agaleus dorsetensis的牙齿有三层珐琅质 , 和晚三叠纪其他软骨鱼类很不相同。Agaleus dorsetensis牙齿齿冠有一对侧齿尖 , 牙齿和长须鲨属(Brachaelurus)及晚白垩纪黎巴嫩及美国的Mesiteia相似 , Agaleus dorsetensis牙齿同时类似石炭纪及二叠纪软骨鱼类Anachronistes及Doratodus。Agaleus dorsetensis牙齿大小比不上纹口鲨及光鳞鲨, 不过Agaleus dorsetensis牙齿要较现存任何须鲨目属种的来得大。从它的牙齿来看 , Agaleus dorsetensis应该是生活于海底并以甲壳类为食的鲨类。
于侏罗纪中期德国及波兰就存活有一种须鲨目成员Folipistrix digitulus , 字首源于folium , 拉丁语意即是叶 , pistrix拉丁语意为鲨鱼 , Folipistrix digitulus也可顺理成章地译作叶齿鲨。叶齿鲨Folipistrix digitulus牙齿非常细小 , 牙齿有较高的齿冠及长有锯齿切边 , 叶齿鲨Folipistrix digitulus牙齿型态类似现存的大白鲨 , 白鳍真鲨及达摩鲨属 , 牙齿没有侧齿尖 , 牙根唇面突起。由于叶齿鲨Folipistrix digitulus牙齿长有锯齿切边 , 它们可能存有和其他须鲨目成员不同的食性 , 可能是一种具掠食性的鲨类 。叶齿鲨Folipistrix digitulus的牙齿于众多须鲨目中是最独特的 , 虽然是归属于须鲨目, 但于归属上还是尚存有疑问的。
须鲨目于晚侏罗纪的欧洲是相当罕有的 , 于德国南部就发现了史前须鲨目成员Phorcynis catulina , Phorcynis catulina最初于法国启莫里阶(Kimmeridgean)地层被发现 , Cappetta于1987年把Crossorhinops minus , Crossorhinops jurassicus 及Palaeocrossorhinus 都划作和Phorcynis catulina属同一物种 , Cappetta于1987年同时把Corysodon及古须鲨属Palaeoscyllium都划作须鲨目 , 但后来于1999年专家根据Corysodon及古须鲨属Palaeoscyllium的牙齿及盾鳞形态而把二者划作真鲨目 , Phorcynis catulina则依然归属于须鲨目。于巴黎,伦敦及慕尼黑都发现了须鲨目的完整骨骼化石 , 这些鲨鱼有较大及圆状的胸鳍及腹鳍 , 有两个几乎相等的三角形背鳍置于身后 , 腹鳍位置于第一背鳍前方 , 有细小及圆状的臀鳍, 臀鳍类似喉须鲛科(Parascylliidae) , 这些鲨鱼的牙齿形态显示和Phorcynis 属关系亲近 , Phorcynis 属于西班牙启莫里阶(Kimmeridgean)地层也有化石记录。
白垩须鲨属种Cretascyllium expansum是被发现于德国森诺曼阶(Cenomanian)地层 , 白垩须鲨属Cretascyllium的分类地位存有疑问 。于英国白垩纪阿尔必阶地层也发现了白垩须鲨属种(Cretascyllium sp)的牙齿化石 , 该化石类似Cretascyllium expansum , 但英国发现的白垩须鲨属种(Cretascyllium sp)牙齿化石有较直的齿冠边及细弱的牙根,侧齿尖也较Cretascyllium expansum的细小得多。英国发现的白垩须鲨属种(Cretascyllium sp)牙齿化石外型呈三角型 ,高度达4.2毫米 , 齿尖是两面凸 , 齿尖长有锯边 , 牙齿齿根长有小孔。
喉须鲛亚目只有喉须鲛科(Parascylliidae)一科, 须鲛亚目则下分六科:鲸鲨科(Rhincodontidae) ,须鲨科(Orectolobidae) ,天竺鲨科(Hemiscylliidae) ,绞口鲨科(Ginglymostomatidae) ,豹纹鲨科(Stegostomatidae) 及长须鲨科(Brachaeluridae) 。大部分属种都是生活于热带,海岸及深海底 , 除了鲸鲨是生活于环球热带地域的远洋。
其他须鲨目成员看似出现的较早 , 长须鲨科(Brachaeluridae)成员出现于早侏罗纪(一亿八千万年前) , 须鲨科(Orectolobidae)出现于侏罗纪中期(一亿六千万年前) , 至于绞口鲨科(Ginglymostomatidae) , 白垩纪早期(一亿五千万年前)便已出现。喉须鲛科(Parascylliidae)牙齿非常细小且不易见,但仍有化石记录 , 于白垩纪至始新世早期喉须鲛科(Parascylliidae)属种中的副鲸鲨属Pararhincodon大部分都保留了牙齿化石 , 但也有完整或部分的椎骨化石发现。
喉须鲛科Parascylliidae一共有三属, 斑鳍鲨属(Parascyllium)一共有四种包括南澳斑鳍鲨(Parascyllium collare) , 澳洲斑鳍鲨(Parascyllium ferrugineum) ,斑鳍鲨(Parascyllium variolatum) 及Parascyllium sparsimaculatum生存于温和的澳洲海岸一带 ; 至于橙黄鲨属(Cirrhoscyllium)中有生存于日本南部的日本橙黄鲨(Cirrhoscyllium japonicum) ,生存于台湾的台湾喉须鲨(Cirrhoscyllium formosanum)及生存于越南Tonkin海湾的喉须鲛(Cirrhoscyllium expolitum) , 橙黄鲨属(Cirrhoscyllium)是生存于接近二百公尺的水深。喉须鲛科Parascylliidae最早于黎巴嫩的白垩纪中期(九千八百万年前)地层有化石记录 , 喉须鲛科(Parascylliidae)化石记录于加利褔亚中新世地层也有化石记录 , 因此喉须鲛科(Parascylliidae)也于特提斯海大幅度扩散 , 现存的喉须鲛属Parascyllium局限于澳洲 , 这可能是发生于澳洲和冈瓦纳古陆 gondwanaland分离后才有的 ,相反橙黄鲨属(Cirrhoscyllium)则局限于亚洲东部。
副鲸鲨属Pararhincodon属种有不少,较著名的包括生活于晚白垩纪土伦阶法国及比利时的Pararhincodon crochardi , 森诺曼阶(Cenomanian)黎巴嫩的Pararhincodon lehmani 及森诺曼阶(Cenomanian)北美的Pararhincodon groessenssi。于德克萨斯州也发现了副鲸鲨属相似种牙齿化石,该相似种cf. PARARHINCODON sp牙齿化石非常细小 ,高度才只有2毫米 , 牙齿有三角型的齿冠及二叶的齿根 , 齿冠基部宽阔 , 有弯曲的末梢 , 牙齿有平滑的锯齿切边 。因为鲸鲨的学名是rhincodon , Para是译作副 , 因此Pararhincodon则被翻译作副鲸鲨属 ,但副鲸鲨属和鲸鲨属体型完全形成强裂的对比 , 副鲸鲨属Pararhincodon是一类体型非常细小的鲨类 , 长度仅只有10厘米以上。
第三纪澳洲及欧洲存有一类喉须鲛科类属的牙齿和现存的大嘴鲨牙齿很相似 ,它们是巨猫鲨属Megascyliorhinus , 被发现的巨猫鲨属Megascyliorhinus化石只有牙齿化石 , 巨猫鲨属Megascyliorhinus有较大向内弯的前齿 , 齿冠舌面存有条纹 , 齿冠唇面凸出 , 平滑及有轻微的条纹。牙齿齿边存有锯边 , 但于齿冠基部则没有锯边存在 。巨猫鲨属Megascyliorhinus牙齿有较高及宽阔的牙根 , 有较深的中央沟槽 。它们也有不对称的侧齿 , 侧齿存有明显的条纹 , 侧齿可能存有小齿尖。巨猫鲨属Megascyliorhinus有始新世英国埃塞克斯的Megascyliorhinus cooperi,渐新世至中新世阿根廷的Megascyliorhinus trelewensis,中新世西欧及上新世北非的Megascyliorhinus miocaenicus , 于比利时渐新世地层也有巨猫鲨属未定种Megascyliorhinus sp的牙齿化石发现。上新世北非发现的中新巨猫鲨Megascyliorhinus miocaenicus牙齿要较中新世的中新巨猫鲨化石样本来得细小 , 但要较始新世的巨猫鲨属种牙齿来得大 。中新巨猫鲨Megascyliorhinus miocaenicus牙齿曾被视作是史前鲸鲨属种,学名更一度被命名作Rhinocodon miocaenicus , 但巨猫鲨属有较细小的牙齿,较直立的齿冠及有锯齿切边,因此后来又被重新划作巨猫鲨属。
长须鲨科(Brachaeluridae)是须鲨科(Orectolobidae)的姐妹群 , 长须鲨科(Brachaeluridae)只有二种包括瓦氏长须鲨(Brachaelurus waddi)及科氏异须鲨(Heteroscyllium colcloughi)。长须鲨科(Brachaeluridae)和须鲨科(Orectolobidae)二个科大多数种类都局限于澳洲新几内亚 , 只有须鲨属(Orectolobus)中的日本须鲨(Orectolobus japonicus)是分布于日本,菲律宾及越南(还有其他须鲨属未定种可能于印尼-马尼拉亚区有分布) , 由于澳洲鲨类的多样性 , 这看似澳洲是长须鲨科(Brachaeluridae)和须鲨科(Orectolobidae)二者世系起源的地方 , 但是欧洲侏罗纪前期至中期的地层也有古长须鲨Paleobrachaelurus的化石记录 ,侏罗纪中期德国的Annea carinata及比利时的Annea maubeaugei 则如谜一样没多少资料可陈述。欧洲侏罗纪中期地层也有史前须鲨科类属Orectoloboides 的牙齿化石记录 , Orectoloboides有不少属种 , 最早的属种是晚侏罗纪英国的Orectoloboides pattersoni , 白垩纪阿尔布阶(Albian)英国东部及法国的Orectoloboides parvulus , 白垩纪阿尔布阶(Albian)法国及德国的Orectoloboides glashoffi , 森诺曼阶(Cenomanian)埃及的Orectoloboides multistriatus ,圣地牙哥始新世地层及澳洲中新世地层也有Orectoloboides 未定种的牙齿化石 , 上新世澳洲南部也有Orectoloboides gipslandicus , Orectoloboides 大概于上新世时期完全灭绝。白垩纪阿尔布阶(Albian)英国东部及法国的Orectoloboides parvulus前齿有幼弱的主齿尖及两个侧齿尖,侧齿齿冠唇面有少量不规则的纵脊。至于现存的妆饰须鲨(Orectolobus ornatus), 于波兰及澳洲上新世地层也有化石记录。史前长须鲨属(Brachaelurus)包括阿尔布阶(Albian)英国及德国的Brachaelurus roklumensis , 坎帕阶(Campanian)美国怀俄明州的Brachaelurus bighornensis , 晚麦斯特里希特阶(Maastrichtian)美国的Brachaelurus hornerstownenesis , 晚麦斯特里希特阶(Maastrichtian)美国达科他州的 Brachaelurus estesi , 于美国达科他州晚白垩纪土伦阶地层也有史前长须鲨属种(Brachaelurus)的化石记录,现存的瓦氏长须鲨于智利及秘鲁上新世地层已化石记录了。 欧洲,北非及西非 , 美国蒙大纳白垩纪前期至始新世前期地层也有其他长须鲨科成员的化石记录 。因此长须鲨科(Brachaeluridae)和须鲨科(Orectolobidae)世系于特提斯海边缘一直有广泛分布但现存却局限于澳洲 , 这暗示二个科大量成员于始新世早期及以后灭绝 , 但只有澳洲的属种却幸存过来。
古长须鲨属Palaeobrachaelurus化石于侏罗纪及白垩纪早期的岩石上都是非常普遍 , 古长须鲨属Palaeobrachaelurus牙齿形态和现存的长尾须鲨属(Hemiscyllium)相似 。古长须鲨属Palaeobrachaelurus属种包括Palaeobrachaelurus mitchelli及Palaeobrachaelurus rocklumensis , Palaeobrachaelurus mitchelli则有较宽阔的侧齿尖及唇面结节 , 牙齿类似须鲨科中的白垩须鲨属种Cretorectolobus doylei及Cretorectolobus gracilis , 白垩须鲨属种中的Cretorectolobus doylei是被发现于英国东部的Speeton Clay Formation , 英国东部阿尔必阶Hunstanton Formation也发现了 Cretorectolobus gracilis的牙齿化石 , 该牙齿化石非常细小 , 高度仅只有3.2毫米。于蒙大纳白垩纪坎帕阶Judith River Formation也发现了Cretorectolobus olsoni及最早的叶须鲨属Eucrossorhinus microcuspidatus的牙齿化石 , 白垩须鲨属Cretorectolobus的牙齿和其他须鲨目成员有明显分别 , 它们有切断食物的齿系。于蒙大纳白垩纪坎普阶Judith River Formation同时也发现了另一类须鲨目成员Cederstroemia triangulate , 该属还有瑞典Kristianstad盆地坎帕阶早期地层的Cederstroemia nilsi 法国坎普阶的Cederstroemia havreensis 。 Cederstroemia 及Cretorectolobus二属一直于美国怀俄明州晚白垩纪的马斯特里赫特期(Maastrichtian)地层也有化石记录, 两个属系可供描述的资料相当稀少。
始豹纹鲨属Eostegostoma仅只有牙齿化石发现 , 它是生活于始新世的比利时 , 最初被认为是属于绞口鲨属(Ginglymostoma) , 因此早斯学名是Ginglymostoma angustum , 但后来又被认为是早期的豹纹鲨科成员 , Cappetta (1987)认为始豹纹鲨属Eostegostoma和长须鲨属(Brachaelurus)关系较近 , 因此又划作长须鲨科,但Compagno (1988)认为始豹纹鲨属Eostegostoma要较短尾铰口鲨(Ginglymostoma brevicaudatum)更像豹纹鲨属的特征 , 因此Compagno把短尾铰口鲨(Ginglymostoma brevicaudatum)划作护士鲨属Pseudoginglymostoma , 但始豹纹鲨属Eostegostoma和护士鲨属Pseudoginglymostoma并非同属。
天竺鲛科(学名Hemiscylliidae)现存只有两个属 , 长尾须鲨属(Hemiscyllium)一共有七种 , 大多分布于澳洲及新几内亚 , 斑点长尾须鲨(Hemiscyllium ocellatum)于所罗门群岛旁的水域也有分布 , 其中的Hemiscyllium galei及Hemiscyllium henryi是生活于印尼一带的新种,二种是于06年被发现。班竹鲨属(Chiloscyllium)有七个地区性的属种各自分布于阿拉伯海,印度及印尼至马尼拉群岛及南日本 , 其中的灰斑竹鲨(Chiloscyllium griseum)同时出没于印尼至马尼拉群岛及澳洲北部 。天竺鲨科是生活于热带水域的鲨类,深海底及浅海沿岸都可发现其踪影 , 体型细小且游动缓慢。
最古老的天竺鲛科化石是来自晚白垩纪(9500万年前)黎巴嫩 , 该鲨类学名是Mesiteia , 其他已知的天竺鲛科化石来自晚白垩纪的德克萨斯州 , 班竹鲨属(Chiloscyllium)出现于晚白垩纪的特立尼达岛, 南达科多,法国,比利时及摩洛哥,始新世前期至中期的多哥; 相反长尾须鲨属(Hemiscyllium)于比利时古新世晚期及始新世前期地层也有化石记录 。
Pseudospinax是一类生存于侏罗纪至白垩纪西欧的天竺鲨科类属 ,属种有侏罗纪卡洛夫阶(Callovian)英国南部的Pseudospinax muftius ,白垩纪坎潘期(Campanian)德国的Pseudospinax hermani , 白垩纪阿尔必阶英国东部的Pseudospinax heterodon , 森诺曼阶(Cenomanian)德国西北部的Pseudospinax pusillus , 德国马斯特里赫特阶(maastrichtian)地层也有未定种Pseudospinax sp的牙齿化石发现。Pseudospinax属种有细小呈三角型的主齿尖 , 有圆状及更细小的侧齿。Pseudospinax属种的牙齿唇面平坦或呈轻微凸起 , 牙齿轮廓像椭圆形,牙齿也长有锯齿切边 ,它们的牙齿也和Protospinax annectans , Protospinax lochensteinensis及长尾须鲨属(Hemiscyllium)的相似。
斑竹鲨属于白垩纪已存在 , Chiloscyllium greeni就是生活于土伦阶时期的西部内海中 , Chiloscyllium greeni牙齿化石被发现于堪萨斯州 , 牙齿化石只有6毫米 , 于英国阿尔必阶地层也发现了相似种Chiloscyllium cf. greeni 的牙齿化石。于古新世晚期及始新世前期班竹鲨属(Chiloscyllium)于特提斯浅海也取得广泛分布且现存属种并分布于印度至西太平洋 , 于比利时的长尾须鲨属(Hemiscyllium)化石却成了谜一般的问题,因为现存的长尾须鲨属是分布于澳洲一带的 , 而比利时的化石样本只有细小的牙齿。于晚白垩纪的西欧已存有长尾须鲨属种, 如英国东部阿尔必阶Hunstanton Formation 已发现了长尾须鲨相似种(cf. Hemiscyllium sp)的牙齿化石 , 牙齿外型和现存的长尾须鲨属种相似,唯一不同的是该长尾须鲨相似种(cf. Hemiscyllium sp)的牙齿化石长有锯齿切边 , 此外于德国森诺曼阶(Cenomanian)至麦斯特里希特阶(Maastrichtian)地层, 摩洛哥及西班牙麦斯特里希特阶(Maastrichtian)至始新世地层都发现了长尾须鲨未定种(Hemiscyllium ssp)的牙齿化石 , 已定种名的史前长尾须鲨属种有坎帕阶(Campanian)至麦斯特里希特阶(Maastrichtian)德国及丹麦的Hemiscyllium hermani , 古新世至始新世中期欧洲(也包括英国)的Hemiscyllium bruxelliensis , 晚古新世英国及始新世比利时的Hemiscyllium dameriesi , 可是史前长尾须鲨属种资料不流通。
于黎巴嫩桑托阶(Santonian)地层发现的斑竹鲨属 Chiloscyllium cheikheliasi 就被归属作新的属阿尔玛须鲨属Almascyllium , 于西班牙北部及法国土伦阶前期地层, 德克萨斯州森诺曼阶(Cenomanian)地层也发现了阿尔玛须鲨属Almascyllium 的牙齿化石。始新世的摩洛哥是鲨类的一片天堂 , 当地也发现了不少鳐类及须鲨目成员的牙齿化石包括原铰口鲨属Protoginglymostoma及须鲨科中的角须鲨属Squatiscyllium牙齿化石, 原铰口鲨属Protoginglymostoma及角须鲨属Squatiscyllium都是属于始新世路德阶(Lutetian),当地也有长尾须鲨属(Hemiscyllium) , Ginglymostoma angolense亲近种(Ginglymostoma aff. Angolense), Chiloscyllium aff. meraense的牙齿化石发现 , 始新世摩洛哥气候和暖, 才能孕育不少软骨鱼类。
长尾须鲨属(Hemiscyllium)能于澳洲水域繁洐辐射, 可能是因为它们的先祖于冈瓦纳大陆分离前已生活于澳洲从而使长尾须鲨属(Hemiscyllium)和班竹鲨属(Chiloscyllium)孤立演化 , 这个假设需要有化石记录支持天竺鲛科是源于晚侏罗纪及早白垩纪。
另外当澳洲漂移和印尼马来半岛相近时容许了天竺鲛科先祖能穿越印尼马来半岛及澳洲间的海洋 , 基于它们的移动性较低 , 因此长尾须鲨属(Hemiscyllium)大约要于渐新世或中新世时期才分布于印尼至马尼拉群岛及澳洲北部这些地区的海洋 , 但长尾须鲨属(Hemiscyllium)及班竹鲨属(Chiloscyllium)于进化枝上的分离要较鲸鲨科及纹口鲨科于古新世时期分离还要来得早 , 因此以上假设没法符合关于长尾须鲨属(Hemiscyllium) 的起源。
光鳞鲨属(Nebrius)于古新世前期已出现 , 现存只有光鳞鲨(Nebrius ferrugineus)一种 , 生活于太平洋及印度洋热带地域的海滨一带 , 于马达拉斯加海域也有发现它的踪影。它们的长度能达3.2米 , 生活于岛架下水深1米至70米 。光鳞鲨(Nebrius ferrugineus)是一种晚上觅食的鲨类 , 以珊瑚虫 , 海胆 , 甲壳类 , 章鱼及鱼类为食。已知的史前光鳞鲨属(Nebrius)属种于古新世前期的欧洲,北美,北非及西非已有分布 , 史前的属种包括N. bequaerti, N. thielensis及 N. blankenhorni 。Nebrius bequaerti生活于古新世时期的西非及摩洛哥 , Nebrius thielensis是生活于始新世中期的比利时及晚始新世的乔治亚州 , Ginglymostoma obliquum是Nebrius thielensis的同物异名,Nebrius blankenhorni 则是生活于始新世中期的非洲。
光鳞鲨(Nebrius ferrugineus)于印度洋围绕的南非至亚洲东南部及澳大利亚都有广泛的分布 , 它也是于太平洋板块中取得广泛分布。绞口鲨属则是须鲨目中唯一生活于大西洋东部及西部,太平洋东部的成员 , 其他须鲨目于那里已灭绝。做成如此分布原因主要是来自因地壳变动产生的山脉和海洋等阻隔 , 绞口鲨属首个化石记录是来自白垩纪前期的立陶宛 , 属种并不少于12种 , 直至中新世时期于美国东部 , 加勒比海 ,西欧 , 非洲西部及北部都已有分布 , 这些史前属种和现存的绞口鲨(Ginglymostoma cirratum)的关系尚待考证 , 而现存的东大西洋,西大西洋及东太平洋的绞口鲨(Ginglymostoma cirratum)相互关系还是尚在研究中 , Castro et al. (2003)近来的研究认为南北美洲绞口鲨(Ginglymostoma cirratum)的数目有别于其他种群水平 , 须鲨目于北美及印尼马来亚的热带亚区的海岸都有高度的多样性及自然地方性。
绞口鲨(Ginglymostoma cirratum)能长达4.3米, 于大西洋及东太平洋的热带温和的水域有其踪影 , 于北美,南美及东非就有铰口鲨的分布。短尾护士鲨 (Pseudoginglymostoma brevicaudatum)则较常见 , 它是栖息于水深12米的鲨类 , 短尾护士鲨于晚上较活跃 , 以无脊椎动物如甲壳类 , 头足类 , 棘皮类 , 软体动物 , 鱼类及黄貂鱼为食 , 短尾护士鲨能长达3米。护士鲨属的化石主要是被发现于摩洛哥晚白垩纪马斯特里赫特期(Maastrichtian)的地层, Pseudoginglymostoma erguitaense 及Pseudoginglymostoma idiri 都是晚白垩纪马斯特里赫特期摩洛哥的史前护士鲨属种 。
铰口鲨属牙齿上有相对较高的中央齿尖及两个侧齿尖 , 而光鳞鲨属(Nebrius)则有相对较低的主齿尖及多倍数的侧齿尖。铰口鲨属(Ginglymostoma)首次于白垩纪马斯特里赫特期(Maastrichtian)的摩洛哥及北美出现 , 晚白垩纪的属种包括摩洛哥的Ginglymostoma. botmaense, Ginglymostoma erramii , Ginglymostoma pectinatum及北美的 Ginglymostoma globidens。北美的 Ginglymostoma globidens牙齿化石被发现于新泽西州 , 特拉华州及切萨皮克 , Ginglymostoma globidens归属是存有疑问 , Ginglymostoma globidens被认为应归属Plicatoscyllium。铰口鲨属(Ginglymostoma)于第三纪的古新世至始新世大放异彩 , 属种于非洲,亚洲及美洲都取得分布 , 包括古新世塔内提阶(Thanetian)尼日尼亚的G. sokotoense , 古新世达宁期(Danian)摩洛哥的G. subafricanum , 古新世塔内提阶(Thanetian)摩洛哥的G. chenanei , 古新世达宁期(Danian)至塔内提阶(Thanetian)摩洛哥的G. khouribgaense , 始新世伊普雷斯阶 (Ypresian)的G. maroccanum及G. maghrebianum,始新世非洲西部及西印度的G. angolense及始新世马里兰州的G. serra。始新世后铰口鲨属种的化石发现便变得稀少了 , 已灭绝的属种包括中新世欧洲及非洲的G. delfortriei及中新世西非的G. malembeense。于北卡罗来纳州Lee Creek Mine的Pungo River formation(早中新世时期)发现了铰口鲨属种的牙齿化石 , 大多牙根都受损 , 但有专家认为这些牙齿化石其实就是属于现存的铰口鲨。
可能是属于现存的铰口鲨的牙齿化石,发现于北卡罗来纳州Lee Creek Mine的Pungo River formation
于第三纪的早期至中期(六千五百万年前至三千五百万年前)软骨鱼类进行了一个前所未有的辐射性进化 , 其中滤食性的软骨鱼类也随之进化 , 包括须鲨目(Orectolobiformes)的鲸鲨 , 鼠鲨目的姥鲨及大嘴鲨 , 鲼形目(Myliobatiformes)的蝠鲼属Mobula及双吻前口蝠鲼(Manta birostris)。这些滤食性软骨鱼类发展至巨大的体型 , 宽阔的大口 , 牙齿变小 , 鳃组织变得精细从而形成过滤微生物 , 这些食性上的急剧转变是什么原因没有任何人知道 。
于美国科罗拉多 , 堪萨斯州及南达科州 , 加拿大萨斯喀彻温省森诺曼阶至土伦阶地层岩石上发现了一种奇特的软骨鱼类的牙齿化石 , 这种史前软骨鱼被命名作Cretomanta Canadensis , Cretomanta canadensis牙齿化石最初于1990年被发现 , 牙齿化石仅只有1.5毫米长 , 牙齿有勾状呈圆锥形的齿冠及厚重的球状齿根 , 齿冠朝舌面弯曲 , 朝齿冠顶部的方向 , 牙齿长有较钝的切边 , 牙根没有营养槽及小孔但具有不规则排列 , 布满血管系统的深坑。由于牙齿实在太细小 ,加上牙齿的独特 , 因此有专家认为Cretomanta canadensis和蝠鱝类或者和鲸鲨科有关系 , Cretomanta canadensis可能只是一种纯滤食的软骨鱼类 , 但归属上仍存有疑问的。
鲸鲨于整个须鲨目是非常独特的 , 因为鲸鲨是有史以来最大的鲨类 , 鲸鲨也是一种大洋性鲨类并有很高的移动性且有环球性的分布。须鲨目(Orectolobiformes)中的鲸鲨科中的古鲸鲨属Palaeorhincodon首次于古新世时期出现 , 于始新世时期已取得世界性分布,于欧洲,非洲,北美及亚洲也发现了古鲸鲨属Palaeorhincodon牙齿化石 。古鲸鲨属包括古新世晚期塔内提阶(Thanetian)的Palaeorhincodon daouii , 始新世早期伊普雷斯阶(Ypresian)的Palaeorhincodon dartevellei及Palaeorhincodon ypresiensis, 始新世路德阶(Lutetian)的Palaeorhincodon wardi , 古鲸鲨属都是生存于比利时,法国及非洲摩洛哥一带,它们的牙齿非常细小,牙齿高度大多才只有3至4毫米 , 牙齿侧部长有小齿尖,牙根形状呈V形 , 牙根中央长有小孔。
现存的鲸鲨生活于热带及气候和暖的海洋 , 会进食浮游生物 , 细小的甲壳类,头足类及小型鱼类 。鲸鲨有较高的移动力,于西大西洋(纽约至巴西中部),东大西洋 (塞内加尔至安哥拉),遍及太平洋及红海,西太平洋(澳洲至日本)及东太平洋(加利褔亚至智利) , 这种大洋性的滤食性鲨鱼于海岸也会被发现 , 甚至会进入礁湖及珊瑚礁。鲸鲨体型一般不超过13米,但也有达18至20米的记录 ,台湾就捕捉过二头长20米及分别重34吨及35.8吨的巨型鲸鲨,也有过21.4米鲸鲨的记录 。鲸鲨上下颌骨各自长有多达300颗的同型齿 , 细小的牙齿只有一个小齿尖。现存的鲸鲨牙齿化石于南卡罗莱纳州渐新世地层,法国及北卡罗莱纳州中新世地层, 马里兰州及维吉尼亚的中新世至上新世地层都有牙齿化石发现。Purdy et al于2001年于Pungo River沉积层也发现了鲸鲨相似种牙齿化石 ,化石是属于中新世时期。牙齿化石非常细小,仅只有6毫米高 , 牙齿侧部下方结实 , 上部的齿冠则显得苗条。齿冠舌面和牙根基部垂直 , 齿尖平滑及长有锯边 , 齿尖且向后下方延伸。牙根是二裂的 , 拉长的牙根侧部压缩 , 有明显的营养孔, 牙根上部有明显的珐琅质覆盖 , 这鲸鲨相似种Rhiniodon cf typus牙齿化石基本和现存的鲸鲨牙齿化石没有多大的分别了。
很少海洋生物有鲸鲨般长寿,就算是雄霸中新世早期至上新世中期的巨齿鲨也没有鲸鲨般万寿无疆 。 鲸鲨自渐新世以来便不断忍受大型鼠鲨类,其他掠食性的鲨类,抹香鲸类及后期兴起的黑鲸类长久威胁, 也要面对像新须鲸类(cetotheres)及其他须鲸类的竞争,但是这些海怪大多已被时间抹灭,只有鲸鲨还巍峨独存。
在本文为化石网论坛精华帖,由网友景德泰岚提供,仅供大家学习参考。
编辑:刘琮滢
2022-12-02
-
称霸一时的节颈鱼目
谈起泥盆纪的盾皮鱼类,那首屈一指的就是其中的节颈鱼目,恐鱼(Dinichthys)和尾骨鱼(Coccosteus)就是代表,它们都属于盾皮鱼亚纲、节颈鱼目,它们生活在泥盆纪时的淡水或滨海环境中,有的发展成为巨型的鱼类
谈起泥盆纪的盾皮鱼类,那首屈一指的就是其中的节颈鱼目,恐鱼(Dinichthys)和尾骨鱼(Coccosteus)就是代表,它们都属于盾皮鱼亚纲、节颈鱼目,它们生活在泥盆纪时的淡水或滨海环境中,有的发展成为巨型的鱼类。其特征是头、胸部被以厚重甲片;头甲和胸甲之间借助一特殊的髁-窝关节相连接,髁突位于胸甲上,髁窝位于头甲上,头甲可以上下活动;眼大而前侧位,眼眶周围有四块巩膜环;胸甲两侧有偶鳍状的骨片或骨棒,不分节不能活动,有背鳍和偶鳍;歪型尾。在大多数状况,上颌有两对骨片,前一对呈锐利的大门齿状,后一对为长切割片状,对应的下颌为一对骨片,其前部呈大门齿状,后部为长切割片状。
尾骨鱼产于英格兰北部和苏格兰中泥盆世的老红砂岩中。该鱼体长22-40cm;头、胸部披重甲,后部则披以皮齿鳞;脊柱的中央椎体未骨化,而脉弧与髓弧均已骨化,并发育良好;歪尾型,尾椎上斜直达上叶末端;有背鳍和偶鳍。
而晚泥盆世的恐鱼(Dinichthys)身体巨大,长可达9m,口中前颌大门齿更为锐利,显系捕食之用,此鱼真可谓当时水中的霸王,早期的鲨鱼,如胸脊鲨、裂口鲨、弓鲛之类,常遭到恐鱼的袭击。前面介绍的邓氏鱼也属于恐鱼家族的大型成员,此外7米长的霸鱼(Titanichthys)和惧鱼(Gorgonichthys)也能与它们分一杯羹,(只是这两类凶猛鱼类的资料目前找到的很少)。
在四川江没和贵州贵阳乌当早泥盆世地层中发现的江油鱼(Kiangyousteus)和贵州鱼(Kueichowlepis)也属此类,因其材料很不完整,尚难复原其形态
在本文为化石网论坛精华帖,由网友父猫提供,仅供大家学习参考。
编辑:刘琮滢
2022-12-02
-
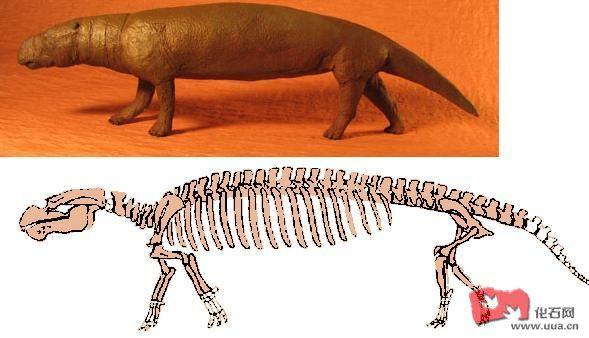
揭开特提斯美人鱼的神秘面纱
美人鱼 , 这名字相信大家也不会陌生了 , 不论于中国 , 日本及西方诸国都会有所流传 , 欧洲传说中人鱼上半身是女性 ,下半身是鱼的身体,伴随着长长的秀发漫游特提斯海。所谓的人鱼 , 其实就是现存的儒
美人鱼 , 这名字相信大家也不会陌生了 , 不论于中国 , 日本及西方诸国都会有所流传 , 欧洲传说中人鱼上半身是女性 ,下半身是鱼的身体,伴随着长长的秀发漫游特提斯海。所谓的人鱼 , 其实就是现存的儒艮及海牛 , 当时的人们很可能看错了,把这些动物看成怪物并加以流传。而这些所谓的美人鱼 , 除了有段名垂千古的传说外 , 背后亦曾有过一段光辉岁月, 以下为本人根据一些连结而整理的资料 , 从而揭开特提斯美人的神秘面纱 !
海牛目跟长鼻目Proboscidea 关系较接近 , Tethytheria 是由海牛目(Sirenia) , 长鼻目(Proboscidea) , 绝种的索齿兽目(Desmostylia)及重脚目(Embrithopoda)组成 。特提斯兽Tethyeria看似是演化出最原始的有蹄哺乳类髁节目 (condylarths) , 出现于特提斯海(古地中海)的海岸 。
根据形态学及分子学研究指出 , 海牛目的祖先跟鲸豚类及鳍脚类等祖先关系遥远 , 尽管大家都是同时于水中生活及进化。海牛目祖先于古新世(6500万年至5400万年前)跟其近亲分道扬镳并迅速重返水中 , 后更扩展至全世界。
海牛目最初进化时 , 它们本身已是素食者 , 依靠海草及水生的被子植物为食。直至现在 , 它们是生活于热带海洋并仍然以被子植物为食 。海牛目拥有较大的肺部 , 沉重的骨骼以助它们于浅水中作停留 。 它们的骨骼是肿胀 , 密集脓厚的 , 特别是肋骨 , 经常可于化石中找到。始新世的海牛目成员 , 跟中生代的哺乳类相似 , 相反跟其它新生代的形成对比 , 前臼齿是有5颗而非4颗 , 齿式是3.1.5.3 。现存的海牛目成员则大多保留了一对獠牙似的上颚门齿 , 前方的牙齿如门齿及犬齿都丧失 , 跟早期的齿式不同。
于印度西北部早始新世的Subathu组发现的Ishatherium subathuensis仅只有臼齿M2化石发现 , 化石有四个主齿尖及呈圆锥形 , Ishatherium subathuensis是否属于海牛目是存有疑问的。此外一些海牛目成员如始新世前期的Paralitherium tarkanyense , 始新世中期匈牙利的Sirenavus hungaricus , 始新世中期欧洲的Anisosiren pannonica , 渐新世哥伦比亚的Lophiodolodus chaparralensis , 中新世斯里兰卡的Miodugong brevicranius及中新世法国的Prohalicore dubaleni的分类地位都存有疑问 , 且缺少具体资料 , 在这贴中不详细描述了。
Prorastomidae科 (始新世中期)留有最古老的海牛目化石纪录 , 是类似古新世至始新世髁节目的两栖四足动物 , 但却变得水生专门化 , 例如有收缩的鼻腔 , 鼻侧的气窦丧失 , 肋骨变得肿胀及密集。最原始的Prorastomids 属种能透过骶骨间的关节支持身体重量以便于陆地上移动。头骨前方及下鄂骨显得不偏斜及横向扁平 , 后肢有较为良好的发展。Prorastomids 拥有较直的喙 , 然而后来的海牛目成员不多不少拥有朝下方的猪鼻状突出物。
Prorastomus 结构上介乎特提斯兽tethytheres 及后期的海牛目成员之间 , 尽管Prorastomus可能不是后期所有海牛目成员的直系祖先 , 但是Prorastomus已明显具备一些已知的海牛目成员特征了 , 包括膨大的喙 , 吻突 , 收缩且扩大的鼻孔及五颗前臼齿 。
Prorastomus sirenoides 生存于始新世中期的牙买加 , 骶骨间的关节缩小 , 但仍能于陆地上承托其身体重量。
Prorastomidae中还有一个未命名的种被发现于牙买加的Chapelton组。
Pezosiren portelli (牙买加) 拥有最古老的海牛目化石纪录 , 这种动物有良好发育的四肢 , 骶骨由多椎骨组成并有较强的骶髂关节连接从而于陆地上支撑其身体重量 , Pezosiren portelli 很可能经常于水中生活 。 Pezosiren portelli 以后的中间种化石显示进化的转变是介乎陆栖及水栖之间 , 海牛目的进化可能类似同时代的原始鲸类 , 透过脊椎延长及骨盆位置同时成了浆状物 , 后肢丧失及尾部变大以作划水游动。
原海牛科Protosirenidae
原海牛科Protosirenidae ,生存于始新世中期 , 第二类重要的早期海牛目成员 ,虽然仍保留了良好发展且短小的后肢 , 但肢间关节不牢固 , 这正可说明了于陆地上其后肢是难以支撑其身体重量。原海牛科Protosirenidae中的原海牛属Protosiren于始新世中期得以大量发展扩散 , 由北非的埃及至南亚的印巴间的特提斯海(古地中海)皆有其分布。透过其头骨及部份骨骼 , 得以知道原海牛属Protosiren 是主要生活于海中并甚少留在陆地上的动物。原海牛属Protosiren除了拥有发展良好且短小的后肢外, 口部还有朝下方的猪鼻状突出物以便于海底进食。下颌联合变宽意味着其植食的选择性缩小 , 其犬齿及门牙缩小并最终丧失(不包括獠牙) 。Prorastomid 及原海牛属Protosirenids 最终于始新世晚期(3800万年前) 灭绝。
原海牛属Protosiren 于海牛目中是与别不同的在于胸椎横切面上拥有较大的钥匙孔形的神经中枢管道 , 肋骨首端一般是软骨而非滑膜关节的 , 并且肋骨是缺乏骨肥厚(pachyostosis)。Protosiren eothene 跟其它原海牛属品种不同之处在于体型细小 , 前胸中枢较Protosiren fraasi的要短10-12% 。 Protosiren eothene椎骨关节部分的肋骨首端至少是滑膜的 , 并拥有良好但相对较肋骨首端细小的肋骨结节。此外 Protosiren eothene颈椎及子宫颈椎骨较后期的原海牛属Protosiren 成员来得长 , Protosiren eothene颅基长可达350mm 。
原海牛属Protosiren (埃及 , 北美 , 欧洲及亚洲 )
1. Protosiren eothene , 生存于早始新世中期的巴基斯坦 。
2. Protosiren fraasi , 生存于早始新世中期的印度 , 埃及及匈牙利。
3. Protosiren sattaensis , 生存于晚始新世中期(3900万年前至4000万年前) 的巴基斯坦。
4. Protosiren smithae , 生存于晚始新世中期的埃及 , 为Protosiren fraasi 直属后代 。
5. Protosiren minima , 生存于始新世中期至晚期的法国。
原海牛科Protosirenidae也有新属种于北卡罗莱纳州的Avon Park , Inglis 及Castle Hayne组有所发现 , 另外有些属种化石可能来自佛罗里达晚始新世的Crystal River组 。
原海牛属Protosiren开始发展出类似现今海牛科成员的骨骼 , 但齿式仍然原始落后 , 被认为分为两个不同的世系 :
儒艮类 : Eotheroides (中始新世)拥有较为轻微弯曲的猪状形吻部及细小的獠牙 , 同时也拥有原始的齿式 。后来于渐新世时期兴起了Halitherium 而这些儒艮科成员拥有较弯曲的猪状吻部及较长的獠牙 。当然现今的儒艮类的吻部更为弯曲 , 獠牙更大。
海牛类 : Sirenotherium (早中新世) , Potamosiren (晚中新世 ) 等类似现今海牛类成员已丧失了部分颊齿 ; 早上新世时期 Ribodon 牙齿则具连续替换的能力。
儒艮科Dugongidae
儒艮科从始新世中期/晚期一直延存至现在 , 儒艮科早期的成员已是完全水生 , 并保留退化的后肢 , 这是相当成功的海牛家族类群并于后来兴起了海牛科 , 渐新世时期(3800万年前至2300万年前)一度是儒艮科的黄金时期 。儒艮科下分Halitheriinae亚科及儒艮亚科Dugonginae , Halitheriinae亚科包括Eotheroides , Prototherium ,始海牛属Eosiren , Halitherium, Metaxytherium 及加勒比海海牛属Caribosiren , 儒艮亚科Dugonginae包括Dioplotherium , Corystosiren 及Rytiodus等于中新世早期均有化石纪录 。
于中新世后(结束于500万年前) , Halitheriinae亚科在加勒比海便消声匿迹 , Halitheriinae亚科有以下属种:
Eotheroides : Eotheroides aegyptiacum 化石被发现于埃及开罗的始新世中期地层 , 稍后于始新世晚期的埃及法扬(Fayum)海床也有所发现 , 于2006年印度发现的新种Eotheroides babiae仅只有下颌及牙齿化石发现。始新世中期北非的Eotheroides属被视作最古老的儒艮科成员 , 前上颌骨有一对细小的獠牙 , Eotheroides属有双脊牙型的臼齿 。Eotheroides属和晚始新世南欧的Prototherium属有密切的关系 , 二者都保留了3.1.5.3 的齿式 。
Prototherium : Prototherium veronense , 生活于始新世晚期的意大利 , 最原始的儒艮科成员之一 , 未成年的齿系相当清楚 , 拥有5颗原始的前臼齿 。过去一直没有未成年的儒艮科齿系化石的发现 , 但是Prototherium veronense 的齿系发现却令人震惊 , 因齿系跟现存的海牛属(Trichechus)的相似 。 此外Prototherium还有Prototherium intermedium被发现。
始海牛属Eosiren , 生存于始新世中期至晚期的地中海 , 有以下属种 :
1.Eosiren abeli , 生存于渐新世早期的埃及
2.Eosiren libyca, 晚始新世
3.Eosiren stromeri
4. Eosiren imenti , 早渐新世的埃及
始海牛属Eosiren大多只有下颌化石 , 众多始海牛属种以伊曼地始海牛Eosiren imenti于海牛目进化史中占最重要的角色。伊曼地始海牛Eosiren imenti的颅基可长达400mm , 从伊曼地始海牛Eosiren imenti发现的地点来看 , 伊曼地始海牛Eosiren imenti有很高的耐盐性 。伊曼地始海牛Eosiren imenti 和晚始新世的Eosiren libyca 及 "Prototherium" intermedium相似 , 于系统分枝学上和Halitherium schinzii 有密切关系 , 伊曼地始海牛Eosiren imenti可能是源于Eosiren libyca 。
伊曼地始海牛Eosiren imenti被认为可能和后来的儒艮亚科Dugonginae,中新海牛亚科Miosireninae及海牛亚科Trichechinae的起源有关 , 儒艮亚科Dugonginae首次于晚渐新世美国东南部出现 , 已知最古老的儒艮亚科属种是Crenatosiren okieni , 此外最古老的海牛科则是晚渐新世的Anomotherium 。
Halitherium出现于晚始新世时期并延存至晚渐新世时期的欧洲 , 并显然于大西洋及加勒比海演化成Metaxytherium 。Halitherium鳍状肢中的趾骨间存有间隙, 后肢退化但趾骨已不存在了 , 不过Halitherium还存有基本的大腿骨连接着退化的骨盆上 , Halitherium 有延长的肋骨增加肺容量以便有效控制浮力 。Halitherium taulannense被发现于2001年 , 生存于晚始新世的法国 , Halitherium taulannense的形态介乎于始新世的海牛目成员及早渐新世欧洲的Halitherium schinzii。
Halitherium属种如下:
1.Halitherium schinzii
2.Halitherium christolii
3.Halitherium antillense
4.Halitherium alleni
5.Halitherium taulannense
加勒比海海牛属Caribosiren只有Caribosiren turneri一种 ,生存于晚渐新世时期的波多黎各(Puerto Rico)。加勒比海海牛属Caribosiren头骨可达300 mm, 吻部偏斜75度及没有獠牙 , 可能专门以海草及根茎为食 。于渐新世结束后再没有加勒比海海牛属Caribosiren的化石记录。
Metaxytherium生存于中新世至上新世 , 跟Crenatosiren及Dioplotherium共存于北佛罗里达 , 跟杜氏海牛属Dusisiren 及 Dioplotherium 共存于墨西哥的下加利福尼亚州 , 分类学上Hesperosiren 跟 Metaxytherium 同义 。Metaxytherium属种有明显向下导向的吻部 , 有细小至中度大小的獠牙 , 以长在水深超过1米的细小海草为食 。
Metaxytherium krahuletzi , 生存于早期中新世的欧洲 ,可能是所有Metaxytherium 成员的直系祖先。
Metaxytherium medium , 生存于欧洲 , Metaxytherium krahuletzi的直系后代。
Metaxytherium serresii , 生存于欧洲 , Metaxytherium medium的直系后代。
于上新世时期 , 其中部分海牛离开地中海 , 它们代表着欧洲至北非间特提斯海中最后的海牛并灭绝于上新世晚期 。于北非利比亚中的Sahabi组中 , 于Metaxytherium serresii 骨骼中发现了一些咬痕 , 显示可能受大型鲨鱼攻击 ,并于Metaxytherium serresii 化石地四周找到巨齿鲨(Carcharodon megalodon)化石 , 意味着当时鲨鱼会捕食海牛目并扮演着重要的生态角色。
Metaxytherium subapenninum(Metaxytherium forestii ) , 生存于欧洲, Metaxytherium serresii的直系后代。
Metaxytherium crataegense(Metaxytherium calvertense/ Metaxytherium riveroi) 全长约3米 ,生存于中新世中期 , 散布于美洲大西洋 , 东太平洋墨西哥的下加利福尼亚州及加利褔尼亚洲一带。
Metaxytherium crataegense 的化石骨架,拥有1500万年之久的历史,于1929年,被发现于佛罗里达州西北部 ,现收藏于佛罗里达州国立大学的地质学研究室。
Metaxytherium arctodites ,分布于旧金山一带 ,跟无齿海牛亚科Hydrodamalinae 有直属关系。Metaxytherium arctodites颅基长可达415mm。
儒艮亚科Dugonginae是儒艮科中唯一生活于上新世时期的加勒比海一带 , 于这时期加勒比海一带的Halitheriinae化石纪录亦消失不见 , 异海牛属Xenosiren(Dioplotherium后代)可能跟Corystosiren一样存活于上新世早期。 晚上新世时期加勒比海地区有一种类似现今儒艮的儒艮科成员 , 但却拥有较大的獠牙(未命名), 它的直系祖先不明 。于晚上新世时期(180万年前) , 儒艮类看似于西大西洋全部灭绝 。 儒艮亚科有演化出较大及刀片状的獠牙趋势 , 对比渐新世至中新世时期Halitheriine那些细小且近似圆锥的獠牙 , 儒艮亚科Dugonginae 更能有效进食海草的根茎。儒艮亚科繁衍于加勒比海至西大西洋地区及进行幅射形进化 , 于晚渐新世至早中新世时期分布于全世界的热带地区 。
Crenatosiren olseni生存于渐新世晚期的美国东南部 , 相比后来的儒艮亚科成员 , Crenatosiren的獠牙比Metaxytherium的要来得大 , 被认为是用作挖掘根茎 。
儒艮(Dugong dugon ) 这是唯一现存的儒艮亚科成员 , 现分布于三十七个国家 , 由非洲西南方和马达拉斯加岛四处的印度洋海岸线 , 包括红海 , 马纳尔湾(印度半岛南端及斯里兰卡之间) , 鲨鱼湾 , 西澳大利亚 , 印太平洋群岛 , 托雷斯海峡(Torres Strait) , 太平洋则南达赫维湾(Hervey)及摩湾(Moreton) , 北达日本冲绳岛。
儒艮的体型大而呈纺锤状,体长约2.4至2.7米,3米以上的个体相当少见,一般而言雌性的体型会比雄性大一点 。皮肤光滑,外观呈褐至暗灰色,腹部颜色较背部来得浅,体表毛发稀疏。颈部短,但仍能有限度的转动头部或点头。前肢短、呈鳍状,末端略圆而缺乏趾甲;胸鳍是幼儒艮主要的推进力来源,成年后则转变为以尾鳍为主。乳房1对,乳头位于前肢基部处。儒艮没有外耳壳,只看得到小小的耳孔,眼睛也很小。鼻孔位于吻部顶端,周围有皮膜可在潜水时盖住鼻孔。宽而扁平的嘴位于厚重吻部的末端下方,嘴边的短须是进食时的重要工具。儒艮有2对门齿,上、下颚各有3对前臼齿与3对臼齿,但所有牙齿不会同时存在,随着年龄增长,它们会失去第1对门齿、所有的前臼齿与第1对臼齿,剩余的两对臼齿则会终生成长。雄性的第2对门齿在12-15岁时会穿透上颚外露,但当闭上嘴时会被厚重的上唇盖住而看不见;雌性极少见到外露的门齿,但少数会保留至30岁左右。
儒艮的肺很大,从胸部一直延伸至肾脏附近,由呈水平的横隔膜将其与其它脏器分隔。支气管深入肺的大部分区域,细部分支少且大半由软骨构成,目前仍不清楚儒艮潜水时肺部是否会和鲸类一般有塌陷的情形。跟其它海洋哺乳动物比起来,它们的脂肪层厚度较薄,同时身体周边似乎不存在热逆流交换系统(heat countercurrent exchange system),肢体末端的动脉与静脉紧邻以维持体热不散失的一种血管构造),这可能与它们栖息于温暖海域有关。胃的构造简单,大肠很大(达胃的两倍重),推测是纤维素的主要消化场所,长度达25米以上,相当于小肠两倍的长度。
一些儒艮亚科成员如Rytiodus, Corystosiren, 异海牛属Xenosiren及Dioplotherium等属有扩大的刀片状獠牙以掘取海草 , Rytiodus及Corystosiren的獠牙比Dioplotherium还要宽阔及更锐利。现存的儒艮也有演化出较大的獠牙 , 但看似只是用作社交。
Dioplotherium属
Dioplotherium manigaulti 分布于晚渐新世的佛罗里达 , 属较分歧的专科并跟Crenatosiren 属同时期的儒艮亚科成员。 Dioplotherium manigaulti及Metaxytherium floridanum头骨均可达460 mm。
Dioplotherium allisoni 于早中新世时分布于巴西 , 加利褔尼亚和墨西哥下加里福尼亚州 ; 中新世中期则分布于阿根廷。于加利福尼亚Dioplotherium allisoni 跟史德拉大海牛的祖先杜氏海牛属Dusisiren 分布有所重叠。Dioplotherium allisoni下颌偏斜度比吻突还要大15至20度 , 牠有着Dioplotherium manigaulti及Rytiodus的特征 , Dioplotherium allisoni被认为是异海牛属Xenosiren的先祖。
异海牛属Xenosiren 仅只有尤卡坦异海牛Xenosiren yucateca , 分布于晚中新世至早上新世的尤卡坦半岛 , 为Dioplotherium allisoni的直属后裔。尤卡坦异海牛Xenosiren yucateca和Corystosiren varguezi同样有着较大的獠牙 , 尤卡坦异海牛Xenosiren yucateca有独特的头骨结构 , 下颌前端有紧密的嚼肌可推动獠牙来挖掘根茎 , 但上颌的牙槽突退化 , 被认为以较细小的根茎进食。Dioplotherium及异海牛属Xenosiren世系于上新世时期完全灭绝。
Corystosiren varguezi分布于上新世早期的尤卡坦半岛及佛罗里达 , 但世系也可来贯及中新世时期的当地 , 独特之处在于拥有非常厚重结实的头盖骨。
Rytiodus capgrandi生存于中新世 , 分布区域跟Dioplotherium allisoni及 Metaxytherium重叠于欧洲 , 北非 及巴西 。 Rytiodus capgrandi长度几乎是现今海牛类的两部 , 全长6米 , 拥有一对鳍状物 , 流线型的身体及尾鳍 。 较平坦的猪状形吻部可容许Rytiodus于较浅的海岸进食 , 不同其它儒艮成员 , Rytiodus capgrandi拥有较短的突出獠牙以榨取沙中的食物。
巴拉特海牛属Bharatisiren一共有两种包括印度巴拉特海牛Bharatisiren indica及喀奇巴拉特海牛Bharatisiren kachchhensis ,印度巴拉特海牛Bharatisiren indica属晚渐新世时期 , 牠的发现显示儒艮亚科是于晚始新世或早渐新世时期越过大西洋进入印非地区 ; 至于喀奇巴拉特海牛Bharatisiren kachchhensis属中新世 , 形态和印度巴拉特海牛Bharatisiren indica的很相似 , 喀奇巴拉特海牛Bharatisiren kachchhensis可能和现在的儒艮属起源相当接近。
于印度也发现了其它儒艮亚科的化石包括早中新世的印度海牛属种Indosiren koeningswaldi 及Domningia sodhae , Indosiren koeningswaldi只有不完整的化石 , 于印度尼西亚也发现了另一属种爪哇印度海牛Indosiren javanense , 属中新世中期 , 印度海牛属Indosiren的数据不流通。
Domningia sodhae被发现于印度喀曲 , 属早中新世 。Domningia sodhae的颅基长可达510 mm , 属体型较大的儒艮亚科成员 。 Domningia sodhae和巴拉特海牛属Bharatisiren前臼齿同样有着多牙根 , 前上颌骨有着较长的鼻突 , 二者的上腭形态都很相似 , 有平坦的剪切型的獠牙。Domningia sodhae , 巴拉特海牛属Bharatisiren及Dioplotherium manigaulti被视作儒艮亚科的基层类群。
侏儒海牛属Nanosiren属于儒艮亚科基层成员 , 一共有二种包括晚中新世委内瑞拉的桑切氏侏儒海牛Nanosiren sanchezi及早上新世佛罗里达的加西亚侏儒海牛Nanosiren garciae , 加西亚侏儒海牛Nanosiren garciae被认为源于桑切氏侏儒海牛Nanosiren sanchezi。加西亚侏儒海牛Nanosiren garciae全长仅只有2米及重达150公斤 。侏儒海牛属Nanosiren有细小的圆锥状獠牙 , 吻部偏斜 , 侏儒海牛属Nanosiren可能以沿岸的二药藻属(Halodule)及喜盐草属(Halophila)等细小的水生植物为食。
除了上述的儒艮属成员 , 还有一种未被命名的新种 , 生存于晚上新世时期的佛罗里达 , 跟现存的印太海牛关系接近 , 最近才被认知的加勒比海及西大西洋地区的儒艮亚科成员。
北太平洋的Metaxytherium 属种于中新世时期进化成无齿海牛亚科Hydrodamalinae 并以史德拉大海牛灭绝作终结 , 无齿海牛属(Hydrodamalis)中最大成员更可长达九米以上。这些种类都能成功适应气候及寒冷的水温 , 并以海藻类为食。无齿海牛属(Hydrodamalis) , 固名思义 , 就是完全没有牙齿 , 并且拥有缩短 , 脚抓似的鳍状肢用来收集植物及避开暗礁 ,同时亦没有趾骨。人类对史德拉大海牛的过渡捕杀使其灭绝于十八世纪。
无齿海牛亚科Hydrodamalinae特征在于新的适应方式 , 跟其它海牛目成员的底部觅食方法背离 , 体型增大 , 扩大领域至寒冷的水域 , 能进一步适应太平洋冰冷的水温并以大型褐藻作食粮取代原本以海草为食。
杜氏海牛属Dusisiren
Dusisiren jordani , 生活于中新世中期(1200万年前至1000万年前)的加尼褔尼亚 , 为 Dusisiren dewana 直属祖先 , 分布区域跟Metaxytherium 及 Dioplotherium 重叠 , 全长4.3米。
Dusisiren dewana , 生活于中新世中期(1000万年前至900万年前)的日本 , 形态学及年代学介乎于杜氏海牛属及无齿海牛属之间 , 全长3至4米。
无齿海牛属Hydrodamalis
Hydrodamalis cuestae是生存于上新世前期(由5百万年至3百万年前) , 分布于加利褔亚及墨西哥的下加利福尼亚州California & Baja 一带。Hydrodamalis cuestae 的头骨中亦是海牛成员中最大的 , 同时亦是有史以来最大的海牛目成员 , 长达10米, 重达22,046磅 , 亦被认为是史德拉大海牛Hydrodamalis gigas的祖先 。
日本北太平洋发现了无齿海牛属Hydrodamalis spissa的化石 , 和杜氏海牛属有一定的相似性。于Dusisiren dewana , Dusisiren takasatensis及 Hydrodamalis spissa , 它们的吻突功能上的发展和史德拉大海牛相似 , 它们颌骨咀嚼轮廓狭窄及矩形 , 翼突腹侧宽大且和翼状肌相连。 Dusisiren dewana及 Dusisiren takasatensis 的夹肌发展类似Dusisiren jordani而不像Hydrodamalis spissa , 半头夹肌的发育则类似史德拉大海牛Hydrodamalis gigas 。Dusisiren takasatensis 有其它特征 , 鳞状骨有较长的S形脊 , 有较大的枕髁及出众的枕蝶骨 , 相对Dusisiren dewana , Dusisiren takasatensis外枕骨有较厚的侧缘。Dusisiren takasatensis 有强健的夹肌 , 半棘肌及头长肌 , 颈部有足够的力量可把头部升降。于无齿海牛亚科中胸椎中部有较低的刺突 , 这意味着轴上肌的发育较少, 椎骨柱灵活性较慢 , 它们游动时像静止一样。
史德拉大海牛Hydrodamalis gigas 于19世纪时曾被归归Rytina 。于1741年在白令海峡发现,发现者为博物学者乔治.斯特拉,当时他正与探险家维他斯.白令一同旅行,而他也是已知唯一看过大海牛活体的生物学家。
史德拉大海牛首次被人类发现时,其分布范围大致位于白令海的司令群岛周边,往东达阿留申群岛西侧,往西至堪察加半岛。由上新世(Pliocene)与更新世(Pleistocene)的无齿海牛属化石记录分析, 过去史德拉大海牛的分布范围可能广达整个北太平洋中北部海岸地带,东侧达下加利福尼亚半岛与现今墨西哥西岸约北纬30度处,西部则到日本的本州岛岛岛(南限约北纬37度)。
当斯特拉第一次描述大海牛的时候,大海牛的群体已经很小,分布地区很有限,数量仅余1,000至2,000头。在人类发现它们后不久,白令海地区拥入许多猎人,这些人的目标是捕杀生存于当地的海獭与北方海狗以获取毛皮,而为了补给需求造成了大海牛的悲剧。这些人大量捕杀大海牛,吃它们的肉、用它们的皮来制造与修补船只。
部分学者认为,海獭的减少可能也是大海牛灭绝的原因之一,在其它生活于白令海的动物中,海獭可能是与它们关系最密切的哺乳动物,因为海獭会捕捉啃食海藻的海胆,如果放任海胆漫无止境的增加,很快地海藻林就会被破坏殆尽。因此当时海獭的减少可能也让大海牛目临食物短缺的压力。
最后一头大海牛纪录是于1768年被杀 , 距离被发现时整整27年便宣告灭绝 。 从史德拉形容所得 , 这巨大草食动物于北太平洋的白令岛及铜岛的数目约为 1500 至 2000 头左右 。这种巨大的动物约长约7. 5米,体重4,500至5,900公斤。 大海牛是没牙齿的 , 嘴里只有两片又平又白的骨头,一块在上颌,一块在下颌 , 两片骨头带有凸起的皱褶以便咀嚼海藻。斯特拉的记录中提到它们会“近岸集体进食,幼兽会被围在群体中央保护” ,而且似乎不会潜水,至少进食时它们只将部分身躯埋入水中,在岸边啃食潮线附近接近海面的大型藻类, 估计其觅食深度不超过1米深 , 单兽甚至可把小艇撞翻。
关于大海牛的文献记录中,曾提到有1头雌性大海牛被捕杀拖上陆地,而另1头与它同行的雄性大海牛就在岸边停留不肯离去,时间至少持续2天以上。加上其它群体观察记录,分析大海牛可能是1夫1妻制,配偶关系可能会维持相当长的时间。斯特拉留下的记录中提到,大海牛多在早春时交配,怀孕期在12个月以 上,终年可生产,不过秋天是分娩的高峰期。事实上斯特拉在白令岛停留的时间从1741年11月底至1742年的8月,由时间上看来他可能不曾亲眼目睹初生 不久的大海牛。
海牛科Trichechidae出现于晚始新世或早渐新世(约3800万年前) ,可能起源于儒艮亚科Dugongidae 。于上新世时期(500万年前至180万年前) , 海牛科Trichechidae的早期成员Ribodon 生活于北卡罗莱纳洲的加勒比海/西大西洋区域一带。于晚上新世时期 , Ribodon 把其活动范围扩展至南美之外。 Ribodon 及其后代海牛属Trichechus 有额外的臼齿 , 它们演化出特殊的牙齿替换系统,以因应食用多纤维食物造成的牙齿磨损。海牛科仅具臼齿, 更新方式不是一颗掉了后再重新长出新牙,而是整列牙齿由颚的末端水平地往前移动,当牙齿移动至颚的最前端时,牙根会逐渐被吸收终至脱落。海牛科Trichechidae可能曾被局限于南美的海河交界及沿海一带直至晚中新世才有所改变。当地它们以内河植物为食 , 而儒艮科则生活于西大西洋及加勒比海并开拓该区的海草牧场。上新世中期(1000万年前至500万年前)安第斯山脉形成时倾倒大量淤泥并把大量营养溶解于南美河流中 , 直接刺激水生的大型植物生长 , 特别是草本植物禾本科(Gramineae)。海牛科成员为了适应新的食物来源 , Ribodon首先拥有额外的臼齿能作水平替换 , 稍后臼齿的尺寸缩小 , 珐琅脊长度增加 , 珐琅褶变得复杂 , 齿排中牙齿数目增加 , 这都于海牛属Trichechus中表现出来。
中新海牛亚科Miosireninae 是海牛亚科Trichechidae 的姐妹组份 , 现已归入海牛科 , 最初海牛科Trichechidae是被认为起源于原海牛科Protosirenidae 及于始新世中期出现的, 但后来中新海牛亚科Miosireninae被认为是根源于儒艮亚科Dugongidae 并兴起于晚始新世或渐新世早期 。
Anomotherium只有Anomotherium langewieschei 一种 , 渐新世时期。
中新海牛属Miosiren生存于中新世中期的欧洲 , 一共有Miosiren kocki 及 Miosiren canhami 两种。Miosiren kocki 全长可达3至4米 , 牠可能是螺食性 , 以獠牙于海底挖出螺类进食。
海牛亚科Trichechinae
第三纪的海牛科成员可能存活于亚马逊盆地三侧的江河一带 , 于晚中新世时期美洲中部海道的出现使得海牛亚科成员得以分布广泛 , 沿着加勒比海及热带的南美大西洋沿岸 , 直到太平洋沿岸皆有其分布 。中新世生活于海河沿岸及泻湖的Metaxytherium 出现并和海牛科产生激裂竞争因此迫使海牛科扩大生境寻找更多的新鲜水生植物 。
Sirenotherium只有Sirenotherium pirabense一种 ,可能是属于中新世早期的海牛类 , 亦可能是属于早期的儒艮类 , 这是1967年Paula Couto 根据巴西找来的两颗牙齿及颅后的节片而描述的。
Potamosiren只 Potamosiren magdalenensis一种 , 生存于中新世中期(1500万年前至1000万年前)的哥伦比亚 , 很可能是属于最早期已知的海牛亚科成员 , 跟Metaxytherium ortegense 同义。Potamosiren magdalenensis是欠缺像后来的海牛科属种具有的额外牙齿能作水平替换这特征 , 这暗示带硅酸的草本植物禾本科(Gramineae)并非其日常食物中最重要的部分。
Ribodon只有Ribodon limbatus 一种 , 生存于晚中新世至早上新世(500万年前至600万年前 ) 的阿根廷及卡罗莱纳州北部 ,可能演化出后来的海牛属Trichechus , 牙齿演化出能无限制性水平替换而这特征于现今海牛类中是存在的。
海牛科为了适应新的食物来源 , Ribodon首先拥有额外的臼齿能作水平替换 , 稍后臼齿的尺寸缩小 , 珐琅脊长度增加 , 珐琅褶变得复杂 , 排齿中牙齿数目增加 , 于海牛属Trichechus中表现出来。
海牛属Trichechus从上新世晚期或更新世(180万年前)兴起并一直延存至现在 , 海牛属Trichechus最初可能分布于多盐分的水域 , 后来发展多样化 , 于第三纪的安第斯山脉形成时扩大其生境进入亚马逊盆地并发展成亚马逊海牛 。 晚上新世时期加勒比海的儒艮类灭绝时 , 西印度海牛则迅速填补其真空地带。
安第斯山脉形成时 , 太平洋的Ribodon就这样进入亚马逊盆地并演化成现今海牛属Trichechus。亚马逊海牛Trichechus inunguis种群发展迅速 , 而且具备特有的特征 , 臼齿变得细小及复合 , 以多种水生与半水生维管束植物为食。趾甲失去 , 倍数染色体数目增加 。出生时身长体约75-85cm、10-15kg , 最大身长体重记录分别为2 . 8m及480kg , 寿命约60-70年 。
西印度海牛(Trichechus manatus ), 出现于130万年前的更新世早期 , 最初出现于佛罗里达 , 更新世晚期至全新世扩展至牙买加 , 路易斯安那州 , 俄亥俄州 , 阿肯色州 , 佛罗里达州 , 南卡罗莱纳州 , 北卡罗莱纳州 , 马里兰及新泽西州等地 。出生时身长体重分别约80至160cm及30kg , 雄性最大身长纪录分别为3 . 7m , 雄性最大身长纪录分别为4 . 1m , 最重可达1,400kg , 寿命约60~70年 。
与其它海牛相同,西印度海牛有流线形的身躯,背脊宽阔而无背鳍。头小,浑身呈灰色,但有些个体外观呈褐、红、或白色,原因可能是藻类或藤壶附着于皮肤表面。皮肤厚而紧实,表面粗糙,体毛稀疏甚至无毛。雌性体型一般较雄性略大。海牛仅具臼齿,会从上、下颚基部水平地往前移动更换。
以下是有关西印度海牛的亚种 :
1. 安地列斯海牛(Antilles Manatee, Trichechus manatus manatus)。
2. 贝克和林海牛 (Trichechus manatus bakerorum ) , 生存于晚更新世 (120,000-125,000年前) 的美国东南部 , 佛罗里达 , 北卡罗莱纳州 , 路易斯安那州等北美地域 。
3. 佛罗里达海牛(Folorida Manatee, Trichechus manatus latirostris)。
4. cf. Trichechus sp. , 一种未定的相似种 , 生存于上新世至更新世时期的巴西亚马逊盆地西部 , 跟佛罗里达海牛Trichechus manatus latirostris相比 ,安地列斯海牛Trichechus manatus manatus较类似这绝种海牛。
西非海牛(Trichechus senegalensis)于晚上新世或更新世时期由南美扩散越过大西洋至非洲 , 现分布于塞内加尔至安哥拉 , 同时西非海牛也是最少被人类研究的现存品种 。出生时身长体重分别约80至150cm及30kg雄性最大身长纪录为3 . 3m、雌性最大身长纪录为3m , 重量最大可达1,400kg , 寿命可能达60年以上。
西非海牛的外观接近其近亲西印度海牛,体型呈纺锤状,背部宽阔无背鳍,头部比例小。体色灰,皮肤厚而硬,略微粗糙,体毛稀疏。栖息于海岸的个体可能有藤壶附着。眼睛小且略微突出(与西度印度海牛不同,其眼睛较为深陷),没有外耳壳。吻部比西印度海牛肥厚而较短,可能是为了适应摄食浮水、挺水植物与生长于浅滩的植物。前肢外侧有三至四个趾甲残留。仅具臼齿,会不断地更换。
侏儒海牛Trichechus bernhardi 是近来被发现于亚马逊河流的海牛属新种 , 牠长度仅只有1.3米 及重60公斤 , 腹部有白色斑点 , 侏儒海牛Trichechus bernhardi 进食的水生植物种类也跟亚马逊海牛Trichechus inunguis 的有所不同 , 此外侏儒海牛Trichechus bernhardi也偏好较浅的急流环境 , 这和亚马逊海牛Trichechus inungui的完全相反 , 许多科学家都认为侏儒海牛Trichechus bernhardi是属于新种 , 但是于世界自然保护联盟却未被确认的。
在本文为化石网论坛精华帖,由网友秋叶月岚提供,仅供大家学习参考。
编辑:刘琮滢
2022-12-02
-
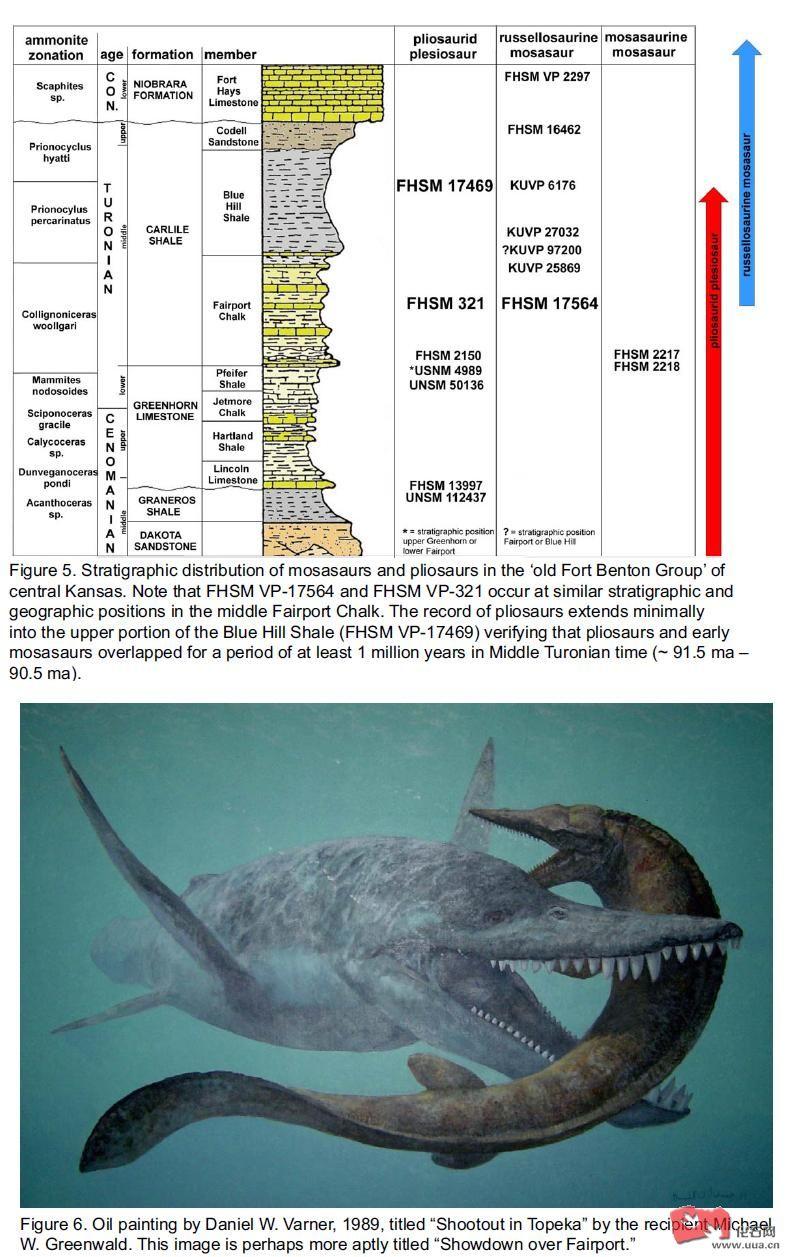
双龙会
沧龙类大概在土伦阶便进入西部内海 , 在堪萨斯州发现的颅骨前额(化石样本KUVP 97200) , 尾椎及躯干椎骨(化石样本FHSM VP2217, FHSM VP2218) 都是属土伦阶 , 化石样本KUVP 97200被认为是属于扁掌龙亚
沧龙类大概在土伦阶便进入西部内海 , 在堪萨斯州发现的颅骨前额(化石样本KUVP 97200) , 尾椎及躯干椎骨(化石样本FHSM VP2217, FHSM VP2218) 都是属土伦阶 , 化石样本KUVP 97200被认为是属于扁掌龙亚科(Plioplatecarpinae) 。 至于化石样本FHSM VP2217只是前尾椎化石 , 椎体侧长约10.3mm 。 化石样本 FHSM VP2218只是躯干中段的椎骨 , 椎体侧长约15.2mm , 由于样本的大小差异 , 两具化石样本被认为不是属同一个体 , 这两具化石样本被认为是归属沧龙亚科(Mosasaurinae) , 牠们比达拉斯龙属(Dallasaurus)还要早。
在堪萨斯州也发现了海王龙亚科(Tylosaurinae)的中段背椎化石 , 化石样本编号为FHSM VP-16462 , 该化石属晚土伦阶中期 。堪萨斯州也发现了另一具海王龙亚科(Tylosaurinae)的化石样本 , 化石样本编号为FHSM VP-2297 , 该化石被确认为海王龙属未定种(Tylosaurus sp) , 全长约6.7米 , 化石样本属早科尼亚克阶(Early Coniacian) , 在当地最常见的海王龙亚科(Tylosaurinae)要数晚科尼亚克阶(Late Coniacian)的堪萨斯海王龙( Tylosaurus kansasensis)。
在新墨西哥州也发现了科尼亚克阶(Coniacian)的海王龙相似属(cf. Tylosaurus sp)的脊柱化石 , 化石样本编号为NMMNH P-22142 , 该脊柱化石全长约160 cm , 估计该个体要比一头7米长的海王龙还要长11% 。在日本北海道穂别也发现了坎帕阶(Campanian)至马斯特里赫特阶(Maastrichtian)的海王龙属未定种(Tylosaurus sp)化石 , 此外穂别当地也发现了同时代的棱沧龙(Mosasaurus prismaticus)牙齿化石 , 棱沧龙(Mosasaurus prismaticus)化石样本展现了异齿性的特征 。以上这些化石记录都显示海王龙亚科(Tylosaurinae)在这段时期迅速崛起并一直向外扩散 , 直至白垩纪结束前仍和沧龙属(Mosasaurus)一争长短。
至于在德克萨斯州发现的所谓森诺曼阶(Cenomanian)沧龙类已被证实为厚根齿鱼属(Pachyrhizodus)的化石(注: 厚根齿鱼属(Pachyrhizodus)和希氏根齿鱼没有任何关系) , 至于土伦阶的沧龙类化石主要被发现在安哥拉 , 哥伦比亚及摩洛哥 , 至于森诺曼阶(Cenomanian)沧龙类化石则被发现在俄罗斯南部 , 而日本北海道穂别也发现了桑托阶(Santonian)或科尼亚克阶(Coniacian)的所谓浮龙属未定种(Plotosaurus sp)的尾椎骨化石 , 对于该所谓的浮龙属未定种(Plotosaurus sp)化石很值得令人怀疑。
以前认为最古老的罗赛尔龙亚科(Russellosaurine)成员是罗塞尔龙属(Russellosaurus), 罗塞尔龙属(Russellosaurus)化石被发现在德克萨斯州 , 化石属早土伦阶中期 , 大概距今9200万年 。此外在堪萨斯州中部也发现了土伦阶沧龙类残骸化石 , 包括四个背椎(化石样本KUVP 6176) , 下颌残骸(化石样本KUVP 25869) , 两个尾椎(化石样本KUVP 27032) , 以上这些化石样本最初被认为和沧龙亚科(Mosasaurinae)有亲缘关系 , 但是近来被认为是罗赛尔龙亚科(Russellosaurine)。
近来在堪萨斯州发现了一具罗赛尔龙亚科(Russellosaurine)化石样本 , 化石样本编号为FHSM VP-17564 , 仅只有七块连续性的尾椎化石 , 化石属早土伦阶中期(Early Middle Turonian) , 化石样本FHSM VP-17564是堪萨斯州发现最古老的罗赛尔龙亚科(Russellosaurine)化石。化石样本FHSM VP-17564七块尾椎合共长约18.7cm , 根据板踝龙属(Platecarpus)及海王龙属(Tylosaurus)的骨骼比例来推算该个体总长约3.2米。不过化石样本FHSM VP-17564这七块尾椎比例上要比板踝龙属(Platecarpus)及海王龙属(Tylosaurus)来得长 , 这显示该个体有相同比例的尾部。牠的尾椎比例上明显比以后的板踝龙属(Platecarpus)及海王龙属(Tylosaurus)要长 , 对海生爬行类而言 , 尾部的骨骼及鳍浆缩短更有利于对海洋环境的适应 , 这暗示早期罗赛尔龙亚科(Russellosaurine)属种对海洋的适应程度还是很低。
在堪萨斯州发现的最晚鱼龙类化石大概是阿尔布阶(Albian)晚期至森诺曼阶(Cenomanian)早期 , 取而代之为双臼椎龙科(Polycotylidae) , 早期的双臼椎龙类(Polycotylids)在阿尔布阶(Albian)和鱼龙类共存 , 以后西部内海便充斥着双臼椎龙类(Polycotylids) , 牠们在土伦阶相当繁盛 , 不过这些双臼椎龙类(Polycotylids)恐怕不是早期沧龙类最大的敌人。
在堪萨斯州发现的双臼椎龙科(Polycotylidae)下颌端部及颈椎化石(化石样本FHSM VP-16869) , 化石可能是属于奥氏长喙龙(Dolichorhynchops osborni), 化石样本FHSM VP-16869可能是堪萨斯州发现的最大双臼椎龙类化石
当早期沧龙类刚刚从陆地向海洋发展 , 海里的上龙类仍然有强大的力量要这批海洋蜥蜴明白何谓实力。多脊齿龙属(Polyptychodon)和短颈龙属(Brachauchenius)就是晚白垩纪初的大型上龙类 , 多脊齿龙属(Polyptychodon)主模式种是森诺曼阶(Cenomanian)至土伦阶(Turonian)的褶齿多脊齿龙(Polyptychodon interruptus) , 褶齿多脊齿龙(Polyptychodon interruptus)化石被发现于南达克州及英国 , 褶齿多脊齿龙(Polyptychodon interruptus)的下颌长度能接近2米 , 体长约10至11米 , 在日本桂沢也发现了褶齿多脊齿龙相似种(Polyptychodon sp cf interruptus)的牙齿化石 , 该化石样本属森诺曼阶(Cenomanian)。
日本发现的褶齿多脊齿龙相似种(Polyptychodon sp cf interruptus)化石及桑托阶(Santonian)或科尼亚克阶(Coniacian)的所谓浮龙属未定种(Plotosaurus sp)的尾椎骨化石
多脊齿龙属(Polyptychodon)另一属种郝氏多脊齿龙(Polyptychodon hudsoni)就是土伦阶北美的上龙类 , 体长约6至7米。 在捷克,瑞士及俄国也发现了多脊齿龙属(Polyptychodon)的残骸化石。
另一具被发现于堪萨斯州的大型上龙头骨化石样本FHSM VP-321是属罗氏短颈龙(Brachauchenius lucasi) , 罗氏短颈龙(Brachauchenius lucasi)是森诺曼阶(Cenomanian)至土伦阶(Turonian)的大型上龙类 , 该属在南美也有化石记录 , 在哥伦比亚就发现了更早的短颈龙属未定种(Brachauchenius sp)几近完整的化石 , 化石属早白垩纪巴列姆阶(Barremian), 该个体颅长约1.2米 , 该化石保留的部分长约3.83米(尾部缺失) , 牠和罗氏短颈龙(Brachauchenius lucasi)所属的时期相距什远。
目前发现的最晚的上龙类化石是堪萨斯州发现的化石样本FHSM VP-17469 , 该化石属晚土伦阶中期 , 化石样本FHSM VP-17469的发现更清楚显示上龙类及沧龙类共存在西部内海至少有1百万年 。有一点是绝对可以肯定的是 , 这些早期的沧龙类和上龙类在土伦阶中期是共存于西部内海的 , 这些向海洋进军的早期沧龙类可能只不过是这些大型上龙类的一碟小菜。
堪萨斯州发现的大型薄片龙类或上龙类的肋骨残骸化石 , 属晚土伦阶中期(Upper Middle Turonian)
在本文为化石网论坛精华帖,由网友耽罗山水提供,仅供大家学习参考。
编辑:刘琮滢
2022-12-02
-

巨型短面熊
巨型短面熊属于眼镜熊亚科(Tremarctinae),它们的近亲是现在的眼镜熊(Tremarctos)和已经灭绝的南美短面熊(Arctotherium),短面熊属包括两个种,倭短面熊(Arctodus pristinus)和巨型短面熊,倭短面熊生活巨型短面熊属于眼镜熊亚科(Tremarctinae),它们的近亲是现在的眼镜熊(Tremarctos)和已经灭绝的南美短面熊(Arctotherium),短面熊属包括两个种,倭短面熊(Arctodus pristinus)和巨型短面熊,倭短面熊生活在中更新世的北美的森林中,头骨相对较纤细,面部较长,牙齿较少,四肢较短,主要以杂食为主,食性大概和现代的北美黑熊差不多。而巨型短面熊有两个亚种,分布在阿拉斯加,加拿大育空,内布拉斯加州,加利福尼亚州和犹他州的育空亚种(A. s. yukonensis)和Rancho La Brea沥青坑发现的较小的巨型亚种(A. s. simus),巨型短面熊是冰河时期北美洲最大的陆上掠食者,雄性比雌性还大20%,育空亚种的肩高1.8米,后肢站立可高达3.3米,体重达600公斤,差不多是棕熊的两倍。而最大的育空亚种后肢站立高度达4.3米,体重达800公斤。短面熊的头骨短宽,而且高高隆起,其宽长之比达到80%,因为没有明显的前额,吻部宽阔,所以看起来不像一般的熊头,这就是它为什么叫短面熊的原因。短面熊的视力很好,眼窝有棕熊的两倍大,颧弓和矢状嵴发达,颚骨靠后,颊骨肌肉发达有很大的咬力,犬齿几乎呈很粗大的三角形,可以帮助它们咬碎骨头。短面熊的体型苗条,可以以笔直而不是类似一般的熊的内八字步伐行走。
那么巨型短面熊怎么掠食呢?过去认为很可能是这样的:在北美洲的大平原上一群群巨大的野牛在吃着草,完全没有注意到一个高大的身影已经在注视着它们中的老弱个体,这是巨型短面熊,它突然冲进野牛群,把其中一头野牛直接扑倒,用巨大的熊掌轻松杀死了自己的猎物,因此巨型短面熊有时被称为“噬牛熊”。
但是根据最近的研究巨型短面熊并不是什么可怕的野牛杀手,因为它的骨骼太脆弱和纤细,这样的结构如果直接和巨大的野牛直接冲撞那么这个家伙多半就要丧命在牛角和牛蹄下了,而且巨型短面熊也很难自己捕食小型的猎物,因为巨型短面熊的骨骼也限制了它的转弯能力,而且速度也不快,追不上泰坦驼,野马等动物,那么这个肉食怪物要吃什么呢?Nancy认为巨型短面熊的速度不快,但是像骆驼一样可以长距离行走,这样就可以在北美洲的大平原上四处寻找腐肉,而且当时众多的肉食动物的猎物也是巨型短面熊抢夺的食物,当一头3米长的巨型短面熊用两条后肢站立高3.3米,这个犹如铁塔一般的体型足以吓得比现在最大的非洲狮子还大25%以上的北美狮,刃齿虎和恐狼狼狈而逃,把猎物让给这个超级强盗。
贴出Arctodus simus和Ursus spelaeus的头骨,注意看Ursus spelaeus突出的前额。
在本文为化石网论坛精华帖,由网友三千院凪提供,仅供大家学习参考。
编辑:刘琮滢
2022-12-02
-
食肉古兽比例尺图片
美国沥青坑的食肉动物,短面熊比北方的小,拟狮比北美北方的大。刃齿虎比佛州的大。欧洲维拉方期的食肉动物:犬属,锯齿虎,豹鬣狗,巨猎豹,巨颏虎,硕鬣狗地中海地区图罗安期的肉食动物群。第二个大熊就是著名的美国沥青坑的食肉动物,短面熊比北方的小,拟狮比北美北方的大。刃齿虎比佛州的大。
欧洲维拉方期的食肉动物:犬属,锯齿虎,豹鬣狗,巨猎豹,巨颏虎,硕鬣狗
地中海地区图罗安期的肉食动物群。
第二个大熊就是著名的印度熊。但是和第四个动物--剑齿虎相比,完全微不足道。剑齿虎不愧是有史以来最大的猫科动物之一。
剑齿虎脚下的是副剑齿虎。看起来小多了
北美洲Hemphilian期的肉食动物群,最后一个就是剑齿虎。还是一如既往的巨大呀。第二个熊是郊熊。
值得一提的是倒数第二个,是著名的巴博剑齿虎。当然不是弗氏的。还是没有剑齿虎大。
还有中间的是上犬,最大的犬科动物就出自上犬属
晚更新世欧洲的动物群,不只是有食肉动物了
有洞狮和披毛犀
1~4楼的图,方格的长度是半米,5楼的图里,方格的长度是一米
更新世的北美洲动物图。其实这些动物也有很多是不共存的。比如说Canis latrans 和Canis lupus ,只有晚更新世的中部平原才有。巨大的美洲虎Panthera onca augusta ,在人类时代的加利福尼亚是有的。但是在沥青坑,确实没有发现。再比如说短面熊类的Arctodus pristinus,是只生活在中更新世时期的,而巨型短面熊Arctodus simus差不多是一直到Arctodus pristinus灭绝了才出现。
在本文为化石网论坛精华帖,由网友绫崎飒提供,仅供大家学习参考。
编辑:刘琮滢
2022-12-02
-

古今大型犬科动物
这次来为大家介绍大型的犬科动物。黄昏犬亚科(Hesperocyoninae)里,最大的属就是奥斯本犬属(Osbornodon),奥斯本犬属里,最大的种类,就是Osbornodon fricki。但是,它的大小实际上也就是和现代的狼差不多大。恐这次来为大家介绍大型的犬科动物。
黄昏犬亚科(Hesperocyoninae)里,最大的属就是奥斯本犬属(Osbornodon),奥斯本犬属里,最大的种类,就是Osbornodon fricki。但是,它的大小实际上也就是和现代的狼差不多大。
恐犬亚科(Borophaginae)里,有一些相当大的动物。例如恐犬属(Borophagus)里的Borophagus dudleyi,颅全长可以达到25厘米。同属的Borophagus diversidens虽然与之相比,颅全长稍小,但牙齿尺寸却相当粗壮
不过,说起恐犬亚科真正的庞然大物,还是要数上犬属的Epicyon haydeni。颅全长可达惊人的34厘米以上。几乎和雄性孟加拉虎相当。按照复原图来推测,它的肩高有90厘米。
当然,上犬属也并非都是如此庞大。Epicyon saevus的颅全长就只有21厘米左右,不过和现代亚洲的狼差不多大。
上犬比例复原图
犬亚科的动物里,北美的狼(Canis lupus)有比较大的个子。颅全长有将近30厘米
生活在更新世的南美洲和北美洲的恐狼(Canis dirus)的个子,比现代的北美大灰狼还要大,颅全长也可以达到30厘米。
除此之外,犬属里还有一些大个子,比如说直隶犬(C。 chiliensis)和德氏犬(C。 teilhardi)。颅全长最大者都可以超过25厘米。此外,龙担的短头犬(C。brevicephalus)的颅全长虽然小于德氏犬,但牙齿却更为粗壮。
除此之外的大的犬科动物,就是异豺(Xenocyon falconeri),颅全长有接近30厘米的样子。
我国早更新世的一些豺类,比如说广西巨猿洞的豺也有比较大的体型,大约和现代的狼差不多大。
除此之外,现代的家犬也可以长到很大。例如高加索狗,体重可以超过100公斤
黄昏犬亚科和恐犬亚科的东西,我是参考的王晓明院士的专著。
狼的测量数据是网上搜来的。
直隶犬是参考<古脊椎动物化石手册>
德氏犬参考邱占祥的<龙担>
巨猿洞的豺,参考PEI的巨猿洞,我按牙齿比例推测的
恐犬的大小比较。
可以看出来,Borophagus dudleyi和Borophagus diversidens是其中最大的了。
我把这两种恐犬的脑袋贴出来。
有兴趣的同志,可以按图上的比例尺量一下,我量的结果是,Borophagus dudleyi也不过25厘米。
再贴一个Epicyon haydeni的脑袋。大家可以按比例尺测量。它的颅全长肯定在30厘米以上
其他的,还有一些大的犬科动物,比如说,Aelurodon taxoides的颅全长大约有27-28厘米。
而其他一些Aelurodon属的动物,或者Carpocyon属的大家伙的脑袋也能有25厘米。
而上犬属的Epicyon aelurodontoides,颅全长能超过28厘米。一些牙齿的大小甚至比Epicyon haydeni还要大。
在本文为化石网论坛精华帖,由网友绫崎飒提供,仅供大家学习参考。
编辑:刘琮滢
2022-12-02
-

白垩纪战斗俱乐部:恐鳄真面目
目录1——恐鳄化石鉴赏-[1]皱鳞恐鳄-[2]西部格兰德恐鳄-[3]海切尔恐鳄2——恐鳄大小-(A)西部格兰德恐鳄:ANMH 3073-[1]头骨重组-[2]头骨对比图-(B)西部海切尔恐鳄:CN 9613——其他-恐鳄咬合力-恐鳄和恐龙
目录
1——恐鳄化石鉴赏
-[1]皱鳞恐鳄
-[2]西部格兰德恐鳄
-[3]海切尔恐鳄
2——恐鳄大小
-(A)西部格兰德恐鳄:ANMH 3073
-[1]头骨重组
-[2]头骨对比图
-(B)西部海切尔恐鳄:CN 961
3——其他
-恐鳄咬合力
-恐鳄和恐龙
-恐鳄种类/分布
感谢以上书籍作者
以及
Colbert/Bird(1954年论作者)
恐鳄属有三个命名种
1903年T.W Stanton和John Bell Hatcher发现,并在1909年被W.J.Holland以Bell Hatcher命名的海切尔恐鳄——deinosuchus. hatcheri
1930-1940年,Robert.T.Bird和Brown在一次原本计划挖掘恐龙的行动中发现后来标号ANMH 3073的标本,标本中的几颗较大的牙齿的根部略大于当时在三层展出的雄性霸王龙ANMH 5027的牙齿;当时命名为Phobosuchus Riograndensis,后来改为deinosuchus Riograndensis,称格兰德恐鳄(格兰德河;Rio Grand River);1954年Robert.T.Bird和当时博物馆研究项目的主负责人Colbert一起发布的有关于格兰德恐鳄的论文
第三种是俗称“东部恐鳄”的deinosuchus Rugosus,Rugosus在拉丁文里面是褶皱的意思,褶皱恐鳄才是正确的翻译。deinosuchus Rugosus在南方也有分布,并非完全在美国的东海岸。
大部分学者认为deinosuchus. hatcheri 和deinosuchus Riograndensis为一种动物
2002年David Schwimmer近一步的提出了deinosuchus Rugosus和deinosuchus Riograndensis也是同种,尽管这观点缺少支持。
和肉食恐龙的对比一下可以发现,中小型成年雄性霸王龙(MOR555)的第19节脊椎的长度大概是140毫米,不包括transverse process的宽度180毫米,最大的一具高棘龙第19节脊椎长度135毫米(相对来说第19节都比第16节要大,16同样比12稍大),格兰德恐鳄不包括transverse process的宽度112毫米,长度128毫米,而海切尔恐鳄第十六节长度140毫米,宽度122毫米
后面提到了,海切尔恐鳄的脊椎最长的一块150毫米,比中小型霸王龙和最大的一具高棘龙的长。而最大的褶皱恐鳄的脊椎长度97毫米(对应海切尔恐鳄的脊椎150毫米)
网络上流传格兰德恐鳄头骨2.2米,这个是没有任何根据的。由于最大的一具格兰德恐鳄的原始化石一直都ANMH保管,就以ANMH最大的重建标本为准(其实2000年前其他博物馆也都是参照这个标本)这个标本是根据古巴鳄和湾鳄联合组建的(并不是百度或者wikipedia上面说主要参照古巴鳄,虽然在1954年的论文里面说用了古巴鳄作参考,但是Bird 1985年的备忘录里面提到实际上主要还是参考一具湾鳄)
第一行为格兰德恐鳄,第二行古巴鳄,第三行湾鳄,第三行美洲鳄
可见吻尖至方骨(头骨最大全长)长度167厘米,而吻尖至枕骨末稍1533厘米
最大下颌联合长度180.5厘米,根本就没有所谓的2米头骨。
至于15米的体长是根据一头印度老湾鳄,头骨身长长度1:9设定的,因为这个湾鳄是个极其罕见的特例,所以这个15.2米已经被认为是卫星了。宽吻鳄的头骨身长比例通常是1:7~1:7.5,而下颌联合长度身长比是1:6
而且即使这具标本也被现代学者认为偏大;而且其他问题也很明显,比如恐鳄属实际上是宽吻鳄不是真鳄,头骨前部过狭窄,后部过长,过宽,牙齿数量太少(13颗,而实际数量20-22颗),而牙齿之间多的距离却有太大(褶皱恐鳄的牙齿之间的距离很紧凑)
ANMH的研究人员最后也意识到了这一点,在1997年博物馆重新装修的时候把ANMH 3073给拆了,把原始化石取出,目前原始化石在W,Langston.Jr 手里,2002年他和他的研究组开始计划重新组装该标本
也就是说这具标本已经作废了
其实早在1997年博物馆重新组装以前就有学者提出ANMH 3073不可靠,并给出了新的,更加精确的重建模型,但是没有被重视而已
比如1973年的版本(绘图者Ron Hirzel,Iordansky)
可以看见恐鳄的牙齿有明显的特化,前部的是穿透性的尖牙,后面的是用来积压和粉碎用的低齿冠牙齿
以上是一具大的头骨就是格兰德恐鳄ANMH 3073,下面则是在Alabama的Mooreville Formation出土的一具恐鳄,实际上就是5楼的那个模型的组成部分,
参考5楼的头骨全长94厘米,不过5楼的那具是电脑合成的(把两个标本拼在一起,迁就的密西西比标本的大小),这具标本标本实际上比5楼的,估计全长113-116厘米左右
如果直接放大:ANMH 3073的全长对应8米的Alabama标本大概只有10.6米,是比较低的计算
当对应摆放好了以后,ANMH 3073和较小的Alabama标本在前颌宽度比例是37:28,假设小标本长度113-116厘米,那么ANMH 3037头骨全长151-154厘米
另外最大的一具完整格兰德恐鳄头骨长度131厘米,目前也在W.Langston手上。
按照1973-1979年的版本,ANMH 3073头骨153厘米,和以Alabam版本重建长度相等,但是较宽
格兰德恐鳄的头骨可能确实比较宽(Langston认为比同长度的褶皱恐鳄要宽一些)
虚线是海切尔恐鳄CM961头骨的极限大小,或者Langston发现的新格兰德恐鳄标本的极限大小
最上面的是修整以后的霸王龙FNMH PR 2081头骨的保守版本,长度143厘米(大版本150厘米,最大的霸王龙头骨MOR 008头骨151-158厘米)
最下面的是已经作废的Colbert和Robert.T.Bird的版本,测量尺一共四把。
那么ANMH 3073估计有多大呢,头骨长度大概1.54米
根据下面5个公式计算
第一个,公式一号:
根据头骨计算长度[1]
Geer的湾鳄公式(1974年)
TBL = -(2.54*4.39) + 7.49(头骨全长cm)
导入头骨长度154cm,得出长度11.42米
根据头骨计算长度[2] Woodward的对比雄性密西西比宽吻鳄的公式(1985年)
Ln(TBL) = 2.132494 + 0.95811(LN(头骨全长cm)) 导入头骨长度148.5cm,
得出长度10.5363米
[3]修改的Woodward公式,两步骤
Ln(TBL) = 2.132494 + 0.95811(LN(0.5头骨全长cm))
2* EXP(LN(TBL))
导入头骨长度154cm,得出长度10.829米,
[4]直接对比密西西比宽吻鳄的两个极大个体,分别头骨对比身长为1:7,1:7.2
得出头骨154cm个体,全场10.78-11.08米,取平均值10.93米
[5]塞诺在2001年使用的湾鳄公式
TL = (7.717 * 头骨长度cm) &#8722; 20.224
导入头骨长度154cm,得出长度11.68米
5个公式平均下来长度11.084米,大概和1989年Ross的计算结果相等(Ross认为ANMH 3073下颌联合长度1805mm是正确的,并以密西西比宽吻鳄头骨下颌联合长度1:6的计算,得出11米)
可以看出下颌联合长度对比全长1:6左右
结论:ANMH 3073放了4米左右的卫星
恐鳄的头骨1.54~1.6米的大小还是相当可观的
下面恐鳄头骨是较大个体,虚线是极限大小
160厘米和霍夫曼沧龙,头骨1.71米的Pilosaurus Brachyspondylus和头骨170厘米的Hainosaurus bernardi(2009年文章里面Unusual death of a Cretaceous giant里面提到根据IRSNB-R23头骨重建的海诺龙bernardi种头骨长度可达170厘米)
当然ANMH 3073也不是最大的恐鳄,CM 961的脊椎骨最长度的一块长度150mm(这块倒不是最宽,最宽的那一块是前面提到的16节,宽度122mm,长度140mm,150mm这一节虽然长,宽度只有110mm),根据对密西西比鳄的研究,脊椎骨长度和体长成线性关系,东部最大的褶皱恐鳄(8米)对应的脊椎骨大概长97+mm,海切尔恐鳄最大长度12米
如果对比格兰德恐鳄,海切尔恐鳄长度大概11~12米左右之间(格兰德恐鳄第12节脊椎骨128mm,海切尔恐鳄第16节脊椎骨140mm,按比例靠后的脊椎稍长)
海切尔恐鳄也未必就比波兰德恐鳄要大,因为Langston宣称可能发现了更大的恐鳄脊椎,据说长度可达12米。
对楼上的补充:Wann Langston, Jr.是著名的权威美国古生物学家,2007年底获得古脊椎生物学会最高荣誉奖,出生于1921年,在1950年和另外一位作者共同编写了高棘龙的建属论文(论文截图,见金闪闪的英雄王的帖子:高棘龙的化石图),目前尚在人世,和Schwimmer认识。所谓的更大的格兰德恐鳄的脊椎是在Big Bend地区发现的,也就是说和ANMH 3073出处相同。
另外密河鳄的脊椎-身长比例是参照了1-3.2米的个体,没有用大个体
最大的恐鳄估计极限12米
体重计算:
[1]湾鳄生长公式:体重=8.7463*(10^-7)*(TL(厘米)^3.25)
导入体长CM961的极限身长12米,得出体重8895kg
评论:鳄鱼是年纪越大,身体越胖,所以用生长公式会高估体重,(12米的湾鳄年龄明显太大)但是考虑到真鳄可能比同年龄/同体长宽吻鳄稍微瘦一点,因此超龄湾鳄计算出的体重还是比较可靠的。可以当作上限使用。比较可靠
[2]直接放大最大密西西比宽吻鳄,2002年Schimmer提到了一条5.84米,并推测体重和6米的湾鳄相等的密河鳄,6米的湾鳄935kg
放大5.84米935kg,个体得出8111.79kg
评论:因为使用的密西西比宽吻鳄是极端个体,用极端个体放大推算海切尔恐鳄的大个体,可靠性高。
放大一个精确计算过的3.76米,237.9kg的野生雄性密西西比宽吻鳄((FEMORAL DIMENSIONS AND BODY SIZE OF ALLIGATOR MISSISSIPPIENSIS: ESTIMATING THE SIZE OF EXTINCT MESOEUCROCODYLIANS,2005年),得出7733.49kg,较大个体推算较大个体,而且这个密西西比的腿骨似乎比较短,大型古鳄类的腿骨基本上比放大的小型鳄短。可靠性高
放大圈养个体4米300kg,得出8100kg,可靠性高。
[3]圈养密西西比宽吻鳄公式
LOG(体重)=3.35LOG(体长)-6.10
导入体长10.5米,体重10500kg
导入体长12米,得出16400kg
评论:可以当成笑料,如果不提出是什么动物,我会以为估算的是虎鲸,或者鲨鱼,没有可靠性
Schimmer拿这个公式作为恐鳄体重8500kg为保守的证据,很显然是瞎扯,难的遇到比2007年Henderson的兽脚类头骨公式还不靠普的。
[4]双步骤圈养湾鳄公式,按照头骨计算 (Webb and Messel,1978年)
SVL (cm) = (3.60 * 头骨长度 (cm)) &#8722; 4.30).
log M (g) = (3.2613 * log SVL (cm)] &#8722; 2.0894).
导入头骨160厘米(恐鳄的极限推测),得出7990kg
评论:比较可靠,原因参考公式[1]
[5]修改版本圈综合计算公式 (专门用于巨鳄的,6-12米,综合了成长年龄等因素)
体重=20*10^(3.35*LOG10(0.33*体长)-6.1)
结果是12.0米的8278kg,可靠
其他书籍里面给的重量:
David Schwimmer认为12米的恐鳄7700-8500kg(其实是他自己没有分清美国短吨和公用吨
Erikson/Brochu提出了格兰德恐鳄最大10米,4500kg,
Ross(1989)提出了格兰德/海切尔恐鳄最大11米,体重5500~6000kg
看得出恐鳄是鼍科的,不是鳄科,而帝王肌鳄甚至不是鳄目的
另外前颌骨已经是拼起来的,所以和前面11楼虽然是一个标本,但是形状不一样。
18楼的公式平均下来12米的恐鳄8319kg,当然这个是按身长/头骨计算,属于偏上限的数值,要大于按腿骨计算的鳄鱼(例如卡氏南方巨兽龙按照1985年Anderson的公式,按腿骨最小周长计算4120kg,按二元腿骨公式计算5600kg,多元腿骨公式计算平均6400-6800kg,腿骨公式上限8800~10791kg,Henderson头骨公式计算13890kg,修改头骨公式9941kg,按身长公式计算大约6600~8100Kg,按放大模型计算则是7962~9131kg)
明天谈一下恐鳄的几个较低下限,毕竟兼听则明
当然,就同兽脚类恐龙一样,巨形鳄鱼的下限的同样可靠性比较低。
Body mass as a function of A, total length; and B, femur length in A. mississippiensis and other crocodylians. Regression lines and 95% prediction intervals are for wild alligators
在现存的密西西比宽吻鳄鱼里面,Log腿骨和Log体重关系是线性的,和头骨vs体重可靠性相等。
因此可以使用腿骨公式计算灭绝的鳄鱼;原先计算史前鳄鱼的大小基本上靠头骨,数值偏大,2005年Farlow设计了腿骨公式
ANMH 7139(前面忘了说了,也是最大的美洲鳄标本,几乎和湾鳄最大头骨差不多大)的头骨,吻尖—方骨长度79厘米,格兰德恐鳄ANMH 3073按照10楼Ron Hirzel,Iordansky给的图片,估计吻尖—方骨 154厘米,大概是美洲鳄的194%。第十二节脊椎骨,ANMH 3073是128mm,而美洲鳄是68mm,是美洲鳄的188%
这头美洲鳄估计全长6米出头,那么按头骨比例,ANMH 3073大约11.7米,按脊椎骨大概11.3米
但是按肩胛骨长度计算,(290/188)*6=9.31米,可见恐鳄的腿是相对比较短的。
格兰德恐鳄长度TMM 43632-1按照头骨(147.5厘米,接近和ANMH 3073)计算身长估计10.64米,
而按照腿骨计算,综合下来长度大约8米左右。上面的腿骨计算的independent variable分别是FL(腿骨长度),Fdw,Fdh,Fpmx,Fpmn,Fc(腿骨最小周长),Ftr.(参考26楼)
可以看出按腿骨宽度和周长计算的值数更较腿骨长度为小,可见恐鳄的腿骨是比较细的。
当然也不是所有巨型鳄鱼的腿骨都偏细,列入下面计算的帝王肌鳄,按照赛诺的说法,大概是最大的帝王鳄的75%,大概8.65-9米和计算出来的长度7.25-9.101米还是比较接近的
按体重计算,10.64米的TMM 4362-1恐鳄,按照单元腿骨长度FL公式计算重量1194kg~2893kg,多元公式平均1724kg,最高值数3458kg,腿骨周长1794kg
那么12米的海切尔恐鳄CM96的体重估计是TMM 4362-1的1.48倍,那么海切尔恐鳄按照腿骨长度FL估算:1766kg~4281kg,按照多元公式最高值数5117.4kg,综合公式平均2551kg,按照2665kg.
帝王肌鳄MNN G102-2按照野生个体(*代表是按完全按照野生个体估算的),那么引用完全用野生个体估算的长度9.1米(见楼上表格,携带*),体重2410kg,塞诺发现的最大帝王肌鳄(MNN 640)大概最可靠的长度估算是11.65米
那么体重大约5100kg左右。
去掉几个完全没有价值的数据
可以确定最大的恐鳄属下限4281kg~5117.4kg,最大的帝王肌鳄估计5100kg。按腿骨计算大概帝王鳄略大于恐鳄
PS:因为恐鳄的活动区域大多在沿海,很可能是海生鳄鱼,因此腿骨相对内河鳄鱼更加细,拿密西西比鳄这样的内河鳄鱼去估算误差会比较大,使用美洲鳄个体相对来说误差会小一些,只不过美洲鳄数量是在太少,很难拿美洲鳄鱼作调查。
而对比一下18楼用头骨计算的体重(7000~9000kg之间)
腿骨公式普遍的偏低(已经举了南巨正模作例子了)
综合一下,恐鳄上限8900kg,下限4281kg。(外加一个平均值下限3500kg,根据TMM 4362-1平均下来2349kg)
最可靠值大概是6000~7500kg之间
下面很多人可能问,最大的鳄鱼到底是海切尔恐鳄CM961,帝王肌鳄(MNN640) ,哈氏肌鳄(sarcosuchus hartii)还是巴西普鲁斯鳄?
(只比最大个体,例如帝王肌鳄和恐鳄里面都有不少6-8米的小家伙,巴西普鲁斯鳄里面两具只要标本)
帝王肌鳄(MNN640)的体长已经不用计算了,大概11-12米之间,CM961大概也是11-12米
前面说了按下限比较,海切尔CM961略小于帝王肌鳄(MNN640),只需要计算帝王鳄的上限体重
至于巴西普鲁斯鳄
巴西普鲁斯鳄的最大个体是DGM 527-R,下颌联合长度估计1750mm,大概头骨参考较小的标本UFAC 1403以及完整的UFAC 1118,长度1485mm,
而哈氏肌鳄在一些不入流的资料力量被渲染成14米,哈氏分布在南美
1a 是哈氏肌鳄:这个下颌骨残片一共是43厘米长,估计右边有牙齿15颗,参考帝王肌鳄的头骨(下图,比例尺50厘米),对应部分是51厘米,看见哈氏肌鳄14米肯定是卫星,因为它无法超过帝王肌鳄。大概体重应该是帝王鳄的60%左右。所以巨鳄争霸中哈氏肌鳄首先被取消竞争资格。
下面是Giant Crocodyliform Sarcosuchus from the Cretaceous of Africa 论文里面帝王肌鳄的比例图
帝王鳄的头骨数据:吻尖至方骨173厘米(本人原来说178厘米,属于测量失误,这次放的很大测的,至于媒体说1.8米或者超过1.8米都是没有依据的)
而论文The Giant Crocodyliform Sarcosuchus from the Cretaceous of Africa 里面给出了吻尖到前枕骨150厘米(如果加上枕骨的10厘米,那么就是160厘米,到方骨全长168厘米,和图片给大小稍微有一些误差)
还有需要注明的就是帝王肌鳄的平均大小也不是11-12米,而是9米左右,帝王肌鳄的复原
巴西普鲁斯鳄的大小
根据头骨计算长度[1]
TBL = -(2.54*4.39) + 7.49(头骨全长cm) Geer的湾鳄公式(1974年)
导入头骨长度148.5cm,得出长度11.01米
根据头骨计算长度[2]
Ln(TBL) = 2.132494 + 0.95811(LN(头骨全长cm)) Woodward的对比雄性密西西比宽吻鳄的共识(1985年)
导入头骨长度148.5cm,得出长度10.1597米
[3]Woodward公式,很简单,两步骤
Ln(TBL) = 2.132494 + 0.95811(LN(0.5头骨全长cm))
2* EXP(LN(TBL))
导入头骨长度148.5cm,得出长度10.46米,
[4]直接对比密西西比宽吻鳄的两个极大个体,分别头骨对比身长为1:7,1:7.2
得出头骨148.5cm个体,全场10.395-10.692米,取平均值10.5435米
[5]塞诺在2001年使用的湾鳄公式
TL = (7.717 * 头骨长度cm) &#8722; 20.224
导入头骨长度148.5cm,得出长度11.25米
[6]对比密西西比宽吻鳄的骨骼
可以看出下颌联合长度接近是全长的1:6,按照1:6计算,DGM 527-R下颌联合长度1.75米,全长10.5米
考虑到普鲁斯鳄是内河鳄,两个湾鳄的公式可靠性较小,那么基本上确定巴西普鲁斯鳄最大个体10.5米
PS:David Schwimmer认为较小的普鲁斯鳄UFAC 1403,体重10吨,David Schwimmer认为较大的DGM 527-R体长可达13.5米,体重能到15吨
Campbell说了11-13米,这些都是很早年的数据,基本上没有什么参考价值(而且宽吻鳄就算12米也不会有10吨)不过11米可能是巴西普鳄年老个体能达到的极限长度。
总的来说普鳄长度比恐鳄小一些。
现在前面给的[1][2][4][5](公式[3]没有价值)个公式分别给出巴西普鲁鳄DGM 527-R(头骨1.485米,全长10.5米)和帝王肌鳄MNN640(头骨1.73米,全长11.5米)的上限体重
[1]DGM 527-R:5763.54kg,MNN640:8079.2kg
[2]DGM 527-R:5187.14kg~5434kg, MNN640:7076~7442kg
[4]DGM 527-R:6250kg, MNN640:8982.7kg
如果直接导入173厘米,得到10327kg,当然结果是无效的,因为要指名一下的是该公式最后仔细看了一下,塞诺针对帝王肌鳄这种头比较长的恒河鳄形态的动物的时候是用头骨前颌骨到枕骨的长度,也就是帝王肌鳄160厘米,这个塞诺已经算过了,是7960kg
PS:体重7960kg则是按12.15米湾鳄推测的,所以塞诺对帝王肌鳄的体重估测不是超过8吨,是最大8吨。
[5]DGM 527-R:5470kg, MNN640: 7248kg
也就是说大概帝王肌鳄最高上限比恐鳄稍低,8982.7kgvs8079.2kg. 而下限帝王肌鳄大于恐鳄,平均则是是海切尔恐鳄CM 961比帝王肌鳄MNN 604重10%以上,但是由于海切尔恐鳄和波兰德恐鳄太破碎,比较完整的恐鳄最大10米左右,而且帝王肌鳄完全长成需要60年,恐鳄则需要80-100岁,所以基本上学术界认为帝王肌鳄略大于恐鳄属成员,而巴西普鲁斯鳄体型最小,虽然其平均个体很可能和帝王肌鳄和格兰德恐鳄相等。
David Schwimmer在书中几次强调了恐鳄比所有的大型兽脚类要重,实在是败笔
[1]首先居然忘了提到棘龙
[2]用兽脚类的腿骨公式计算的结果去比较恐鳄的头骨和生长公式,那肯定是恐鳄大了
实际上比最大的海切尔恐鳄大的兽脚类标本(不包括镰刀龙科这些可能是食素的)至少有,括号里面是标本数量
葡萄牙蛮龙(1),霸王龙(7),摩洛哥棘龙(1),埃及棘龙(这个大太多了,都没有必要比),南方巨兽龙(2),玫瑰马普龙(2),撒哈拉鲨齿龙(1)
和CM961差不多大的有纣魁龙(1)。
现在比较一小大小:)*代表不可靠。
按12米(上限长度)的CM 961头骨公式和建模法(放大宽吻鳄等于建模)计算的结果是7700~8900kg之间
用腿骨计算的极限下限是3500~5200kg之间(去掉那些太低的,根本就是一看就知道是胡扯的)
南方巨兽龙
兽脚类用腿骨计算(举一个最著名的标本+一个最大标本)*代表不可靠。
MUCPv-Ch1(12.5米):4200~7200kg(腿骨长度);13980kg*(头骨);建模6650~8800kg
MUCPv-95(13.5米):5200~8900kg(腿骨,放大小的那个南巨);16500kg*(头骨,参考小的南巨);建模,9000~11500kg
霸王龙
FNMH-PR 2081(12.5米):5600~8900kg(腿骨);9100~10500kg*(头骨);建模,10200~11200kg
MOR 008(13.3米):6900~11000kg(腿骨,放大CM 9380);12500kg*(头骨);建模,11000~14000kg
埃及棘龙
MSNM V4047 (16-19米):9000(按腿骨公式计算的似鳄龙为准)~9800kg, 建模,17000~24000kg,公式推测(16000~26500kg*)
撒哈拉鲨齿龙 (鲨齿龙的头骨不止1.6米,应该是1.8米,这个金闪闪的英雄王已经解释过了,塞诺的重建模型方骨有错误)
SGM-Din 1(13.4米):5100~8200kg(腿骨,参考正模);16500kg*(头骨);建模,9000~11200kg
蛮龙(13.4米):4800~7600kg(腿骨/放大北美标本);13000kg*(头骨);建模,8500~10500kg
玫瑰马普龙
MCF-PVPH-108.185(推测13.9~14.2米);建模,10000~12000kg
MCF-PVPH-108.145(推测13.5~13.9米);建模,9000~11000kg
当然恐鳄出现的时候,它是北美有史以来最大的肉食动物(霸王龙在所有恐鳄属已知成员灭绝以后才出现)
在恐鳄比在它之前的最大掠食者伊潘龙和高棘龙大
伊潘龙太碎,没办法估计,但是可以给个范围,大概是按腿骨3000~4500kg,按建模法大概5500~7000-7300?kg(上限),比恐鳄轻至少1500kg
高棘龙两具标本NCSM 14345/ OMNH 10147估计都差不多大,按腿骨周长2400kg,按建模3D大概5500~6500kg,头骨估算5300kg,腿骨估算比恐鳄小,其他的估算也比恐鳄小。
所以说该是谁大,就是谁大
大部分恐鳄的分布限制于Campanian.而Peedee formation和Black Creek 都有一些恐鳄分布
但是不排除这些恐鳄实际上很可能是另外一种生态地位接近现代恒河长吻食鱼鳄的7-8米的大型鳄鱼Thoracosaurus neocesariensis
Schwimmer提出了三种恐鳄实际上可能是一种得观点
以及生态分析,美国内海东西两岸的动物种群很相似(不知名的霸王龙超科是霸王龙形兽脚亚目Appalachiosaurus montgomeriensis,当时哈没有命名 )
当然这只代表一种说法,并非主流
1:霸王龙(10200~14000kg)
2:蛇发女妖龙(1000kg)
3:惧龙CM 9401(5000~6000kg)
4:伊潘龙(5000~6000kg)
5:高棘龙(5300~6200kg)
6:被褶皱恐鳄袭击的肉食恐龙大小(唯一一起可靠记录)
7:东部褶皱恐鳄
8:最大的海切尔恐鳄CM961 (6500-8800kg)
(忽视那个测量尺)
分析褶皱恐鳄的咬合力
整个龟壳厚度(~2厘米),牙齿穿透深度1厘米,估计是比较尖的后部短牙(恐鳄后部牙齿也有尖的,但是发现的后部牙齿30%都是受到压损,甚至短掉的,而前部牙齿15%受到压损)
那么如何根据这个咬痕计算恐鳄的咬合力呢?
第一恐鳄的牙齿和霸王龙的前面的差不多,都是圆锥状的牙齿,可以用计算霸王龙牙痕的方法计算恐鳄咬力
大概就是霸王龙一颗牙齿穿透了11.5mm的骨头,真正造成主要穿透难度的是硬骨层,按figure3,硬骨层的厚度和需要穿透的力量成线形关系。霸王龙的牙齿穿透了2.5mm的硬骨层,需要6400N,假设这颗牙是上颌骨第7颗牙,那么由于机械消耗,实际需要力量7890N,如果是上颌骨第四颗牙,需要10300N, 后来基本上可以肯定是第四颗牙了,高速穿透需要更大的力量+30%,肌肉组织消耗了10%左右的力量,那么总共用掉了13400N的力量。要计算恐鳄的这个咬痕,首先要确定硬骨层的厚度
45楼给出了公式
2305.405(高密度骨骼厚度:毫米)+646.634
那么Bothremys 的龟壳硬骨层的厚度是多少呢?
可以看出D就是Bothremys baberi,和恐鳄咬的那个是同一种动物,ECO就是硬骨层的厚度,按比例2厘米的龟壳,硬骨层大概是小于等于3.4mm
褶皱恐鳄咬Bothremys baberi的分析,另外一个霸王龙,还有一个是西部格兰德恐鳄,最小面的头骨上面的箭头表示牙齿可能的位置(霸王龙其实已经可以确定是靠前面的那颗了)
根据公式2305.405(高密度骨骼厚度:毫米)+646.634计算出需要大约9333.5N的力量,+10%的机械消耗+30%的穿透速度,(由于是直接咬龟壳,所有没有能量浪费在穿透肌肉上),最后估计是13347N
和霸王龙的一颗牙差不多,当然霸王龙的是feeding bite,也就是吃东西时用的力,这条褶皱恐鳄估计是killing bite。当然这些计算都不是很可靠,显然没有参考是否有多颗牙齿同时咬如猎物,或者是下面的牙齿又没有同时发力(controlateralness)
格兰德恐鳄最深的一个牙印大概是在脊椎骨上面穿透了5mm,往高处算,也就是假设5mm都是硬骨层的话,那么至少需要15000N的咬力。这个也是feeding bite
当然这不是指恐鳄拥有超过T-rex的咬力,霸王龙的clashing bite咬力可以穿透3-4厘米的脊椎骨
而格兰德恐鳄的clashing bite记录反倒不这么出色,很浅的咬痕。可能是恐鳄实际上希望的是溺死猎物吧。
恐鳄和同时期恐龙的关系;作者YY恐鳄袭击埃尔伯塔龙,这个吗,看看图,且不说是真是假,估计是12米CM961这个大小的恐鳄袭击亚4-5米长成年埃尔伯塔龙,否则如果是成年埃尔伯塔龙按这个比例恐鳄是卫星
另外就是格兰德或者海切尔恐鳄没有袭击埃尔伯塔龙的证据,而褶皱恐鳄体型比埃尔伯塔龙小(埃尔伯塔龙1620~2800kg),褶皱恐鳄(最大2400kg)而且褶皱恐鳄和埃尔伯塔龙也不重叠。恐鳄袭击恐龙的主要证据是(可能)杀死了一只亚成年的兽脚类,Appalachiosaurus montgomeriensis 大概兽脚类300-400kg,恐鳄个体1000~2000kg
这个比例,咬死了理所当然,咬不死才出问题了呢……
在本文为化石网论坛精华帖,由网友鸿穹神皇上帝提供,仅供大家学习参考。
编辑:刘琮滢
2022-12-02
-
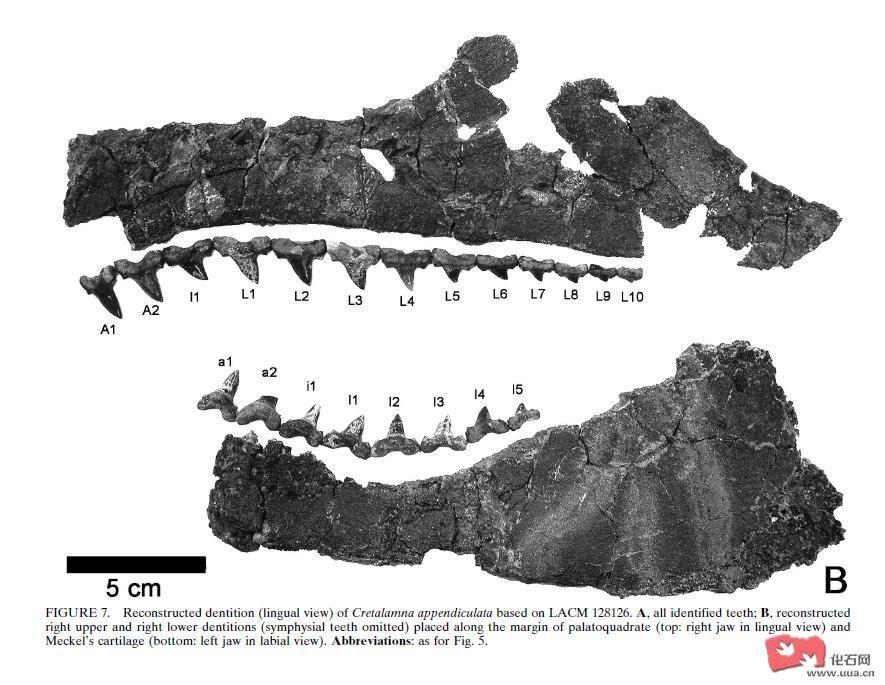
蛇颈龙化石上惊现鲨鱼牙齿 曾遭七头鲨围攻
蛇颈龙化石上惊现鲨鱼牙齿 曾遭七头鲨围攻美国科学家在一具拥有8500万年历史的蛇颈龙骨头化石上发现了鲨鱼的牙齿,经过仔细分析研究后证明,这头巨大的蛇颈龙曾遭到七头 Cretalamna appendiculata(现代大白鲨
蛇颈龙化石上惊现鲨鱼牙齿 曾遭七头鲨围攻
美国科学家在一具拥有8500万年历史的蛇颈龙骨头化石上发现了鲨鱼的牙齿,经过仔细分析研究后证明,这头巨大的蛇颈龙曾遭到七头 Cretalamna appendiculata(现代大白鲨的早期亲属)鲨鱼的围攻,这一重大研究成果将发表在下周召开的第69届古脊椎动物学年会上。
这具被鲨鱼攻击的蛇颈龙化石从日本中部出土,并被安放在东京的日本自然科学国家博物馆内。美国芝加哥市德保罗大学大学副教授Kenshu Shimada表示:“这是鲨鱼吞噬古脊椎动物的一项无可非议的证据。
Kenshu Shimada和他领导的研究小组发现,五颗Cretalamna appendiculata鲨鱼的牙齿镶嵌在蛇颈龙四块不同的骨头上,同时出土的还有同种鲨鱼的80个相关牙齿。牙齿的大小和形状表明,这些围攻蛇颈龙的鲨鱼都是年轻或者年壮的鲨鱼。
Kenshu Shimada依据出土现场留下的各种材料推测出了8500万年前一场惊心动魄的血腥事件的部分过程:“这头蛇颈龙原本居住在浅海区域,后来由于某种原因死亡了,它的尸体漂浮到深海区域,沉入了海洋的底部并形成了今天的化石。它死亡的情形很可能是这样的:至少有六到七头 Cretalamna appendiculata鲨鱼发现了蛇颈龙,并发起了围捕,吞噬它的肉。但现在还不能确定,这头蛇颈龙是立即被吃掉的,还是在很长一段时间内才被吃掉。蛇颈龙如果真如猜测所言受到了攻击,那么这一来自Cretalamna appendiculata鲨鱼的攻击肯定是致命的,因为蛇颈龙骨头化石上显示,其骨头伤口没有任何痊愈的痕迹。”
Kenshu Shimada自幼喜欢查阅和了解有关蛇颈龙和鲨鱼牙齿的资料,在此期间增长了很多的知识。他说:“我感到很有趣,同时充满自豪,这一30年后的发现赐给了我了解远古鲨鱼的牙齿的机会。”
新墨西哥州自然历史科学博物馆馆长托马斯·威廉姆斯(Thomas Williamson)说:“这不是一件关于鲨鱼吞噬蛇颈龙的普通证据,而是Cretalamna appendiculata鲨鱼吞噬蛇颈龙的证据。真正让人感兴趣的是如此多的Cretalamna appendiculata鲨鱼一起合作攻击或分享蛇颈龙。这种捕食方法和现代鲨鱼猎食很相近。”对于蛇颈龙在遭到攻击后的结果,威廉姆斯馆长表示不能排除蛇颈龙成功逃离围堵的可能。
Cretalamna appendiculata
Sept. 17, 2009 - Remains of a shark-bitten, 85-million-year-old plesiosaur reveal that around seven sharks likely consumed the enormous dinosaur-era marine reptile in a feeding frenzy, leaving some of their shark teeth stuck in the plesiosaur's bones, according to a new study.
The findings, which will be presented at next week's 69th Annual Meeting of the Society of Vertebrate Paleontology, are the first direct evidence of the diet and feeding behavior of Cretalamna appendiculata, a now-extinct early relative of today's great white sharks.
The study, which has also been accepted for publication in the Journal of Vertebrate Paleontology, further represents what lead author Kenshu Shimada describes as "arguably the most spectacular case of shark feeding on a vertebrate carcass reported to date."
Shimada is an associate professor at Chicago's DePaul University and research associate in paleontology at the Sternberg Museum of Natural History. He and colleagues Takanobu Tsuihiji, Tamaki Sato and Yoshikazu Hasegawa analyzed the shark-decimated plesiosaur, Futabasaurus suzukii, which was unearthed in central Japan and then housed at the National Museum of Nature and Science in Tokyo.
They found five C. appendiculata teeth embedded in four different bones of the plesiosaur, and additionally discovered 80 associated teeth of this same shark species with the remains. The size and shape of the teeth indicate they belonged to both juvenile and adult sharks.
Based on the physical evidence, Shimada and his team determined what likely happened to the over 30-foot-long dinosaur-like marine reptile.
"The plesiosaur inhabited the near shore, shallow sea," he told Discovery News. "Whatever the cause of its death, the plesiosaur carcass came to rest belly-side up on the bottom of the sea floor, below the reach of surface waves, where mud mixed with sand grains accumulated relatively rapidly."
"Prior to its decomposition, at least six or seven Cretalamna appendiculata individuals, possibly ranging in size from about 5 to 14 feet in length, began to scavenge the plesiosaur throughout its body. Whether or not the feeding activity took place continuously or intermittently is uncertain."
The researchers also haven't ruled out that the huge marine reptile was attacked and killed by the sharks.
"If the plesiosaur had been attacked by one or more individuals of C. appendiculata, it must have been a fatal attack because bones of the plesiosaur immediately around the embedded teeth do not show any indication of bone healing," the scientists concluded.
Thomas Williamson, curator of paleontology at the New Mexico Museum of Natural History and Science in Albuquerque, told Discovery News that the new paper isn't the first reported evidence of shark feeding on a plesiosaur, but it is "the first evidence of feeding by the shark Cretalamna appendiculata."
"It is interesting that such a large number of individuals of C. appendiculata - six or seven - may have participated in the predation or scavenging," Williamson said, adding that "similar numbers of modern shark species are known to behave this way."
Williamson isn't, however, convinced by the inferred position of the plesiosaur during the feeding, as he said it could have moved during the course of the shark feasting.
Shimada said he grew up reading about the plesiosaur in Japanese children's science books, and vividly remembers seeing photographs of the shark teeth in a pictorial science encyclopedia.
"It's rather amusing - and I'm honored at the same time - that I was given an opportunity to formally describe the very same shark teeth about 30 years later," he said.
To find out more about sharks, visit the Shark Weekend website.
Cretolamna appendiculata普遍被认为是大白鲨 , 长肢灰鲭鲨 , 短肢灰鲭鲨及巨齿鲨等的共同祖先
C. appendiculata 可达3米
The C. appendiculata individual likely measured at least 2.3 m and possibly as large as 3 m in total length. The extant phylogenetic bracket approach indicates that C. appendiculata possessed a dental bulla in each jaw. Cretalamna appendiculata was probably an ecological generalist.
在本文为化石网论坛精华帖,由网友宝源崇德提供,仅供大家学习参考。
编辑:刘琮滢
2022-12-02
-

浅谈日本的掠食性抹香鲸类
谢氏虎鲸(Orca semseyi)抹香鲸总科在第三纪有过大量属种 , 目前已知最古老的成员是晚渐新世阿塞拜疆的凯氏法勒西鲸(Ferecetotherium kelloggi) , 已知早期的抹香鲸类还有匈牙利发现的「谢氏虎鲸(Orca sem
谢氏虎鲸(Orca semseyi)
抹香鲸总科在第三纪有过大量属种 , 目前已知最古老的成员是晚渐新世阿塞拜疆的凯氏法勒西鲸(Ferecetotherium kelloggi) , 已知早期的抹香鲸类还有匈牙利发现的「谢氏虎鲸(Orca semseyi)」, 阿根廷发现的普氏蒂普罗鲸(Diaphorocetus poucheti)及巴塔哥尼亚拟艾多鲸(Idiorophus patagonicus) , 北意大利发现的伯尔桑拟艾多鲸(Idiorophus bolzanensis) 。 在中新世中期抹香鲸类大幅度分化 , 在南美 , 北美东部及西部 , 欧洲西部 , 地中海地区 , 新西兰 , 澳洲及日本都有化石记录 , 只有少量化石是属晚中新世至更新世的 , 但是化石保存不完整。以众多史前抹香鲸类来说 , 要数以下的种类最重要 , 在介绍主角之前 , 先简单介绍一下以下重要的角色。
凯氏法勒西鲸(Ferecetotherium kelloggi)
凯氏法勒西鲸(Ferecetotherium kelloggi)体长并不超过6至7米(一说为5米) , 凯氏法勒西鲸(Ferecetotherium kelloggi)有较低而短的头骨 , 横向拉长的头盖骨 , 上枕骨前倾及接近三角型 , 有短而宽的鳞状颧突 , 凯氏法勒西鲸(Ferecetotherium kelloggi)有着异型齿 , 牠的颌骨有多达27颗牙齿 , 下颌中部的牙齿要较其它的大 , 牠的牙齿呈圆锥型并有细小的齿冠及单牙根 。凯氏法勒西鲸(Ferecetotherium kelloggi)虽是目前已知最古老的抹香鲸类 , 但是骨骼上和古鲸亚目却很相似 , 只是颌骨结构有所不同 , 比如齿冠严重退化 , 牙根略微陷入颌骨 , 此外凯氏法勒西鲸(Ferecetotherium kelloggi)骨骼上却也有须鲸类的特征 。
巴罗拉颧突抹香鲸(Zygophyseter varolai)
巴罗拉颧突抹香鲸(Zygophyseter varolai)被发现于意大利 , 属晚中新世中期(距今1100万年前) , 颅骨长约1.5米 , 有几乎完整的骨骼化石被发现 。巴罗拉颧突抹香鲸(Zygophyseter varolai) 上颌骨每边各有13颗牙齿 , 下颌每边各有14颗牙齿 , 最大的牙齿直径占颅长的3.8% , 颞窝长度为颅长的28% , 巴罗拉颧突抹香鲸(Zygophyseter varolai)体长约6.5米至7.5米。
恐齿弓抹香鲸(Acrophyseter deinodon)
恐齿弓抹香鲸(Acrophyseter deinodon)化石被发现在秘鲁 , 化石属晚中新世(600万年前) , 恐齿弓抹香鲸(Acrophyseter deinodon)颅骨长约90厘米 , 上颌每边各有12颗牙齿 , 下颌每边各有13颗牙齿 , 最大的牙齿直径为颅长的3.5% , 颞窝长度为颅长的37% , 从恐齿弓抹香鲸(Acrophyseter deinodon)的头骨长度判断体长约3.9米至4.3米。恐齿弓抹香鲸(Acrophyseter deinodon)及巴罗拉颧突抹香鲸(Zygophyseter varolai)的吻端呈管状 , 吻端的圆锥型牙齿平伏并相互锁合 , 能有效抓捕猎物 , 下颌后列的牙齿横向性平坦 , 能把猎物切成碎块。恐齿弓抹香鲸(Acrophyseter deinodon)及巴罗拉颧突抹香鲸(Zygophyseter varolai)的进食方式大概和古鲸亚目及鲨齿鲸科等具异型齿的齿鲸类相似 , 古鲸亚目及鲨齿鲸类同样有着圆锥型及锋利的前牙及犬齿 , 臼齿及前臼齿呈三角型且横向平坦。
弓抹香鲸属未定种(Acrophyseter sp)
在秘鲁发现的弓抹香鲸属未定种(Acrophyseter sp)属晚中新世(1200万年前至1100万年前) , 颅骨和恐齿弓抹香鲸(Acrophyseter deinodon)的相似 , 不过牠的颅骨要较恐齿弓抹香鲸(Acrophyseter deinodon)的略大 , 吻部也较宽 。 该化石的鳞状颧突及颞窝也和恐齿弓抹香鲸(Acrophyseter deinodon)的有所不同。
麦尔利维坦鲸(Leviathan melvillei)
麦尔利维坦鲸(Leviathan melvillei)也是被发现在秘鲁 , 化石属中新世中期(1300万年前至1200万年前) 。麦尔利维坦鲸(Leviathan melvillei)颅骨长约3米 , 上颌每边有9颗牙齿 , 下颌每边有11颗牙齿 。 麦尔利维坦鲸(Leviathan melvillei)最大的牙齿直径为颅长的4.1% , 颞窝长度为颅长的25% , 麦尔利维坦鲸(Leviathan melvillei)体长约13.5米(以抹香鲸颅宽来算)至17.5米(以巴罗拉颧突抹香鲸(Zygophyseter varolai) 颅宽来算)。麦尔利维坦鲸(Leviathan melvillei)吻背前沿轮廓呈圆状 , 颌骨有非常粗状的圆锥型牙齿 , 这些牙齿能重创猎物及从猎物身上撕开肉块 , 麦尔利维坦鲸(Leviathan melvillei)的进食方式也许和逆戟鲸 , 伪虎鲸及侏虎鲸等相似。
在意大利Astigiana地区发现了三种鲸类化石包括上新贝拉尔迪喙鲸(Berardiopsis pliocaenus) , 索氏抹兽鲸(Physotherium solterii)及原型锯齿抹香鲸(Priscophyseter typus) , 化石属上新世时期。其中索氏抹兽鲸(Physotherium solterii)及原型锯齿抹香鲸(Priscophyseter typus)属抹香鲸类 , 牠们属疑问学名 。索氏抹兽鲸(Physotherium solterii)只有一点颌骨及牙齿化石发现 , 原型锯齿抹香鲸(Priscophyseter typus)化石也太残破 , 难以和其它抹香鲸类作比较从而判断是否属于掠食性抹香鲸类。
除上述的抹香鲸类是具掠食性外 , 还有尖角斯卡尔鲸(Scaldicetus oxymycterus)及四贺噬抹香鲸(Brygmophyseter shigensis)也同样是掠食性抹香鲸类 , 两者同样被发现在日本 。 中新世中期至晚期是中小型须鲸类蓬勃的时期 , 由于食物的丰裕程度使这批掠食性抹香鲸类得以大辐度发展。
四贺噬抹香鲸(Brygmophyseter shigensis)属中新世中期 , 过去的学名包括四贺斯卡尔鲸(Scaldicetus shigensis)及四贺长野鲸(Naganocetus shigensis) 。四贺噬抹香鲸(Brygmophyseter shigensis)的左右下颌齿骨都近乎完整 , 保留的左下颌齿骨长度约120 cm , 根据下颌齿骨长度估算 , 四贺噬抹香鲸(Brygmophyseter shigensis)的颅骨至少长约150 cm , 四贺噬抹香鲸(Brygmophyseter shigensis)体长约7米。四贺噬抹香鲸(Brygmophyseter shigensis)是抹香鲸类演化史中最重要的一环 , 牠有渐新世齿鲸类原始的型态 , 但是同样也有许多抹香鲸类的洐征 , 和四贺噬抹香鲸(Brygmophyseter shigensis)共存的鳄形奥巴斯托鲸(Orycterocetus crocodilinus)及莫理斯管状鲸(Aulophyseter morricei)在抹香鲸类演化史中还只是相对原始的阶段。
四贺噬抹香鲸(Brygmophyseter shigensis)颅骨有非常原始的特征比如倾斜的枕骨盾(occipital shield) , 现存的抹香鲸有垂直的枕骨盾 。四贺噬抹香鲸(Brygmophyseter shigensis)下颌有较大的颞窝 , 这显示所在位置是附有强而有力的颞肌 , 四贺噬抹香鲸(Brygmophyseter shigensis) 颧骨突起部分有较大的鳞状骨 , 鳞状骨侧面膨大 , 四贺噬抹香鲸(Brygmophyseter shigensis)的下颌联合也比现存抹香鲸的短 , 这些都被视作原始特征。四贺噬抹香鲸(Brygmophyseter shigensis)上下颌都长有牙齿 , 珐琅质厚达1mm , 现存的抹香鲸上颌只有退化的小齿龈 , 牙齿是没有珐琅质的 。 四贺噬抹香鲸(Brygmophyseter shigensis)每排颌骨的牙齿数目大约只有12颗 , 牙齿数目和古鲸亚目的相当接近 , 至于现存的抹香鲸下颌的牙齿数目能多达24颗甚至更多 。
四贺噬抹香鲸(Brygmophyseter shigensis)有较大呈卵形的下颌孔(也是声纳的证据) , 四贺噬抹香鲸(Brygmophyseter shigensis)的骨盆(supracranial basin )比抹香鲸(Physeter macrocephalus), 鳄形奥巴斯托鲸(Orycterocetus crocodilinus)及莫理斯管状鲸(Aulophyseter morricei)的要浅 , 四贺噬抹香鲸(Brygmophyseter shigensis)的中筛骨(mesethmoid)形成隔壁把左右鼻骨都分隔 , 左鼻骨孔比右鼻骨孔要大得多 , 四贺噬抹香鲸(Brygmophyseter shigensis)的右鼻骨孔比例上甚至比抹香鲸(Physeter macrocephalus), 鳄形奥巴斯托鲸(Orycterocetus crocodilinus)及莫理斯管状鲸(Aulophyseter morricei)的要细小 , 四贺噬抹香鲸(Brygmophyseter shigensis)右鼻骨孔大辐退化也是一个洐征 , 右上颌骨的后鼻骨位置并没有和颅骨中线的左上颌部分相汇。
四贺噬抹香鲸(Brygmophyseter shigensis)的眶部相对其它抹香鲸类要置前得多 , 牠的眶部位处颧弓前方。四贺噬抹香鲸(Brygmophyseter shigensis)的鼓泡相对其它抹香鲸类来说是很细小的 , 牠的鼓泡保留了古鲸亚目和须鲸亚目的特征。四贺噬抹香鲸(Brygmophyseter shigensis)耳外骨有非常大的乳突(mastoid process) , 大小是莫理斯管状鲸(Aulophyseter morricei)的1.5倍。
四贺噬抹香鲸(Brygmophyseter shigensis)有相对较短的肱部 , 牠的肱部仍保留了和莫理斯管状鲸(Aulophyseter morricei)同样的原始特征 , 四贺噬抹香鲸(Brygmophyseter shigensis)的肱部比凯氏法勒西鲸(Ferecetotherium kelloggi)短而宽 , 凯氏法勒西鲸(Ferecetotherium kelloggi)有修长的肱部 , 这是相当原始的特征 , 不过凯氏法勒西鲸(Ferecetotherium kelloggi)的尺骨和现存的抹香鲸同样有一个刻面 , 这显示凯氏法勒西鲸(Ferecetotherium kelloggi)也同样具有洐征 , 四贺噬抹香鲸(Brygmophyseter shigensis)尺骨有两个刻面 , 现存的抹香鲸已没有这特征。四贺噬抹香鲸(Brygmophyseter shigensis)尺骨未梢也有着凯氏法勒西鲸(Ferecetotherium kelloggi)的特征。
在日本也发现了许多四贺噬抹香鲸(Brygmophyseter shigensis)近亲的化石 , 比如在日本古口(Furukuchi)发现了晚中新世(约600万年前至550万年前)的斯卡尔鲸属未定种(Scaldicetus sp)的前牙化石(化石样本编号为YPM7923) , 化石样本YPM7923有较大的单牙根 , 圆锥型的齿冠 , 齿冠有较薄及具条纹的珐琅质 , 牙根呈长柱状 , 从化石样本YPM7923来看牠和四贺噬抹香鲸(Brygmophyseter shigensis)有非常密切的关系。
群马发现的化石样本GMNH-PV-581属晚中新世中期(距今1150万年前) , 仅只是颌骨残骸化石 , 化石样本GMNH-PV-581保留的牙齿属于颌骨后列牙齿 , 最大的牙齿长约139.6mm。化石样本GMNH-PV-581牙齿间隔宽约11mm至14mm , 四贺噬抹香鲸(Brygmophyseter shigensis)的牙齿间隔宽约11mm至24mm , 两者的牙齿间隔数值不同 , 因此化石样本GMNH-PV-581被认为和四贺噬抹香鲸(Brygmophyseter shigensis)没有任何关系 , 从化石样本GMNH-PV-581齿冠端部磨损程度来看该化石可能属于老年个体 。
茨木市(ibaraki)发现的化石样本INM-4-012885包括吻部残骸及右鼓泡化石 , 化石属晚中新世中期至中新世晚期 , 最大的牙齿长约158 mm。化石样本INM-4-012885鼓泡尺寸和四贺噬抹香鲸(Brygmophyseter shigensis)的相近 , 化石样本INM-4-012885可能是属未成熟的个体。化石样本INM-4-012885也有前列牙齿化石发现 , 牙齿尺寸及弯曲程度和化石样本YPM7923的相近 , 两个化石样本的牙根直径大致相近 , 化石样本INM-4-012885的齿冠比化石样本YPM7923的细小得多 。化石样本GMNH-PV-581及INM-4-012885牙齿形态及尺寸和四贺噬抹香鲸(Brygmophyseter shigensis)的牙齿相近 , 两具化石样本都被视作斯卡尔鲸相似属种(cf . Scaldicetus sp)。
在日本水户市(Mito)也发现了中新世中期的掠食性抹香鲸类的下颌骨及牙齿化石(化石样本NSM-PV 2219) , 化石样本NSM-PV 2219牙齿型态和化石样本INM-4-012885的相似 , 两者的发现地相当接近 , 并且属同一时期 , 化石样本NSM-PV 2219是另一个在日本发现的类似斯卡尔鲸属 (Scaldicetus)的掠食性抹香鲸类化石。
在本文为化石网论坛精华帖,由网友耽罗山水提供,仅供大家学习参考。
编辑:刘琮滢
2022-12-02

