鬣齿兽科(Hyaenodontidae)的一些简单补充
鬣齿兽类(hyaenodontids)是古近纪旧大陆及新大陆占优势地位的掠食动物,属于Creodonta,旗下大约有50个属。包括了4个亚科:原灵猫亚科(Proviverrinae),这是最为原始的鬣齿兽类动物;Limnocyoninae亚科最初现鬣齿兽类(hyaenodontids)是古近纪旧大陆及新大陆占优势地位的掠食动物,属于Creodonta,旗下大约有50个属。包括了4个亚科:原灵猫亚科(Proviverrinae),这是最为原始的鬣齿兽类动物;Limnocyoninae亚科最初现于古新世最晚期的亚洲,是Prolimnocyon属。同食肉类大相径庭的是它们的裂齿并非P4,m1;而是M2,m3。
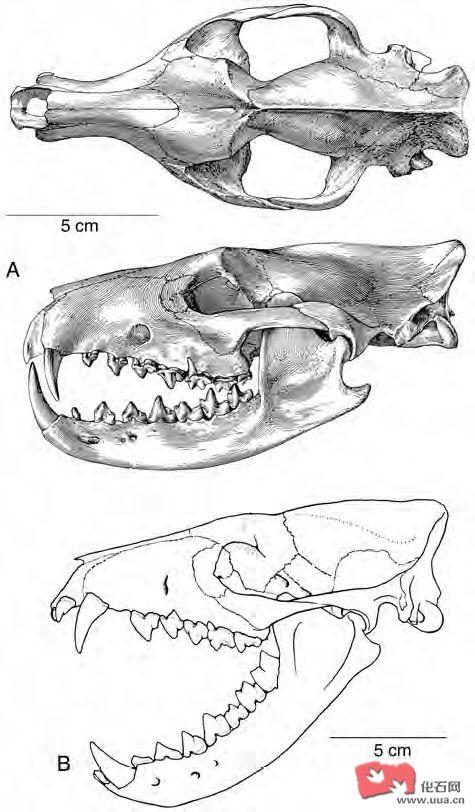
典型鬣齿兽类hyaenodontids的头骨:
A.Sinopa
B.Hyaenodon
这是Prolimnocyon atavus的骨骼重塑
Hyaenodon crusians的头骨
Limnocyoninae亚科的多数成员拥有着短而宽的头骨,这比起鬣齿兽类,倒是更像oxyaenids。Prolimnocyon生活于Wasatchian阶,其颅后骨表明或许适宜于攀爬。
还有个是Pterodontinae,这个亚科有些学者认为
应该和Proviverrinae共同并入众所周知的鬣型兽亚科(Hyaenodontinae)。
下图2个,第一个是Prolimnocyon;第二个就是似鬣狗翼齿兽。
还有一张是我的好朋友Mauricio Anton老师画的Pterodon dasyuroides
提到鬣齿兽亚科,我想大家应该都很耳熟能详了吧。其中的鬣齿兽属(Hyaenodon),是从晚始新世到早渐新世,北美洲,欧亚大陆的优势顶级掠食者。鬣齿兽长有粗壮的前臼齿及特化的M2,m3(鬣齿兽的裂齿),且也没有下后尖。
除了我曾经介绍过的大型鬣齿兽科动物,像是北非的Megistotherium osteothlastes,古北区的Hyainailouros sulzeri,和北美洲的Hemipsalodon grandis,O.M.S.I No.619的个体的颅骨全长甚至达到了骇人的450MM。
而在鬣齿兽属里,Hyaenodon horridus恐怕是目前最著名的种了。它们的脑袋很大,颅骨长度能够达到250-350MM,而整个鬣齿兽属里体型最大的莫过于Hyaenodon megaloides了,其Skull Length 足有400MM,是非常大的食肉动物,在Chadronian阶,Orellan阶及Whitneyan阶的动物群里是典型的顶级掠食者。不过鬣齿兽属里大多数种的体型并不大,像北美洲早Arikareean阶的Hyaenodon brevirostris就是很小的食肉动物。还有如发现于蒙古早渐新世的Hyaenodon chunkhtiensis也是种较小的掠食动物。当然了,蒙古和北美洲都是有大型,甚至可以说是巨型的鬣齿兽种的,比如Hyaenodon pervagus,其m1有一简化的下跟座。
最后说说BBC《与野兽同行》里出现过的Hyaenodon gigas,其实这个种的材料简直少得可怜,就一些前臼齿及几块上颌骨碎片罢了,究竟有多大谁也无从知晓,但是可以肯定这个种体型非常大。我曾经比较过一个Hyaenodon gigas相似种的P2,P3和p3长度,宽度,可以发现这几颗前臼齿比其他的鬣齿兽类大的多的多。在蒙古中部的Taatsiin Gol地区还发现过cf. H. gigas的右爪骨,特别大,比北美洲Hyaenodon horridus的爪子还要大出约50%。
这个就是那块大爪子化石。
还有Hyaenodon gigas相似种的部分前臼齿长宽测量数据。大家有兴趣可以比较看看。
图1:Hyaenodon gigas的正模标本PST 27-10就是一块左上颌骨碎片,上面有P4–M2,由Dashzeveg描述。
图2:Hyaenodon gigas的部分前臼齿。
图3,图4:BBC版本的Hyaenodon gigas。
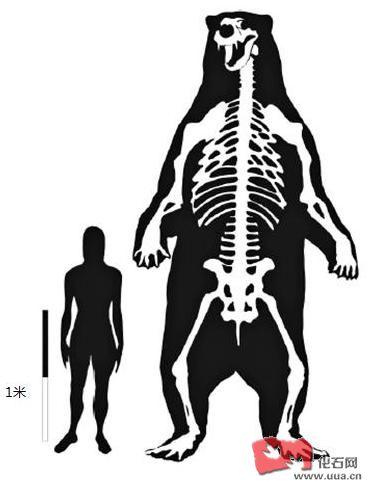
补充一张家犬和Hyaenodon horridus的骨骼对比图,可以看出Hyaenodon的脑袋比犬科动物大的多。
家犬和Hyaenodon horridus的骨骼对比
补充一个Hyaenodon的图,m3有68.8MM
在本文为化石网论坛精华帖,由网友超剑虎提供,仅供大家学习参考。
编辑:刘琮滢
2022-12-02